

Effect of environmental factors on CPUE of shrimp trawls along the west coast of Madagascar based on Bayesian networks
-
摘要: 研究渔业资源与环境因子的关系,并了解种群分布对环境变化的响应机制,是养护渔业资源、实现渔业可持续发展的基础。渔业资源丰度和种群分布受多种环境因素影响,但目前的研究更多关注环境因素的直接影响,较少考虑环境因素间的相互作用。为了探索不同环境因素对马达加斯加西海岸虾类资源量的影响机制与路径,本研究使用2014−2020年该海域捕虾拖网数据,采用贝叶斯网络分析了降水、径流等海洋环境因子与3种主捕虾类单位捕捞努力量渔获量(catch per unite effort,CPUE)之间的网络关系,探索在多种环境因子影响下3种虾类CPUE的潜在驱动因素。研究结果表明:降水、径流、海面高度距平(sea surface height anomaly,SSHA)和海表面温度(sea surface temperature,SST)是影响印度白虾CPUE的主要因素,径流、SSHA、 SST和叶绿素a质量浓度(chlorophyll a concentration,Chla)是影响独角新对虾和短沟对虾的CPUE的主要因素;降水通过不同路径影响其他环境因子进而对3种虾类CPUE产生间接影响:降水通过径流、SST和SSHA的途径对印度白虾产生间接影响,但对于独角新对虾和短沟对虾,降水通过径流、SST、SSHA和Chla的途径产生间接影响。研究结果揭示了马达加斯加西海岸降水和其他海洋环境因子不仅对3种虾类CPUE产生直接影响,降水还能通过影响其他环境因子对虾类种群资源变动的间接影响途径和机制。Abstract: Investigating the relationship between fishery resources and environmental factors, along with understanding species distribution response mechanisms to environmental changes, provides fundamental insights for fisheries conservation and sustainable management. While both resource abundance and species distribution are influenced by multiple environmental factors, existing research has primarily emphasized direct environmental effects, with insufficient attention to inter-factor interactions. This study examines the mechanisms through which diverse environmental factors affect shrimp resources along Madagascar's western coast, utilizing shrimp trawl fishery data (2014−2020) and Bayesian network analysis to investigate relationships between precipitation, runoff, marine environmental factors, and catch per unit effort (CPUE) of three key shrimp species. Our analysis identified critical drivers of CPUE variation under combined environmental influences. Results demonstrated that precipitation, runoff, sea surface height anomaly (SSHA), and sea surface temperature (SST) predominantly influenced Fenneropenaeus indicus CPUE. For Metapenaeus monoceros and Penaeus semisulcatus, runoff, SSHA, SST, and chlorophylla concentration (Chla) constituted primary controlling factors. Precipitation exerted indirect effects on all species' CPUE through environmental mediators: impacting F. indicus via runoff-SST-SSHA pathways, while influencing M. monoceros and P. semisulcatus through runoff-SST-SSHA-Chla interactions. These findings clarify both direct effects of precipitation and marine environmental factors on shrimp CPUE, and reveal cascading indirect impacts where precipitation modulates population dynamics through environmental intermediaries, elucidating pathway mechanisms underlying these ecological relationships.
-
Key words:
- bayesian networks /
- environmental factors /
- shrimp /
- CPUE /
- precipitation /
- runoff
-
1. 引言
马达加斯加西海岸海域底部地形平坦,渔场分布在多个内陆河流与海水交汇的区域,内陆河流携带大量的有机物入海,形成富含营养物质的环境,为渔场提供了丰富的饵料和栖息地[1]。该地区的捕虾拖网渔业始于20世纪50年代[2],并于1967年开始商业化拖网捕捞,作业渔船数量从1970年19艘逐渐发展至1996年的峰值77艘,随后逐渐减少至2020年的71艘[1]。然而,渔业的持续发展对主要捕捞种群,如印度白虾(Fenneropenaeus indicus)、独角新对虾(Metapenaeus monoceros)和短沟对虾(Penaeus semisulcatus)等[3]造成了较大压力。随着渔业的平均捕捞量和单位捕捞努力量渔获量 (catch per unite effort,CPUE)出现波动与下降,以及渔业管理部门采取限制准入许可、设立禁渔期等措施,表明该海域的虾类资源已处于过度开发状态[4−5]。
环境因子是影响近海虾类种群分布及资源补充的关键因素。目前,国内外相关研究多集中于海洋物理和生物因素,海表面温度(SST)被认为是影响虾类生长、繁殖和洄游等行为的重要影响因素,是表征虾类栖息地环境的关键指标[6];海面高度距平(SSHA)提供了栖息地状况,并对虾类资源的丰度和空间分布产生影响[7];叶绿素a质量浓度(Chla)通过影响饵料生物的分布,间接作用于渔业资源的空间格局[6]。此外,淡水补充[如降水(Precipitation)和径流(Runoff)]对沿海及河口渔业资源具有显著影响[8−9],改变了海洋初级生产力、养分流动及生物群落组成,进而影响渔业资源的丰度[10−11]。尽管已有研究探索了淡水补充对虾类资源丰度的影响[7, 12−13],但针对马达加斯加西海岸的相关研究较少,特别是淡水补充对近岸虾类资源的影响。因此,深入研究淡水补充对近海虾类资源的影响,掌握马达加斯加西海岸主要虾类种群栖息分布及其与环境因子的关系对于养护虾类资源、实现渔业可持续发展具有重要的意义。
随着渔业资源影响因素研究方法的发展,广义线性模型(Generalized Linear Model,GLM)[14]和广义加性模型(Generalized Additive Model,GAM)[15]等方法被广泛应用于环境因子与物种资源丰度之间的关系研究。然而,这些方法通常只关注相互独立变量及其交互作用对种群丰度的直接影响,而忽略了因子之间的因果关系及其可能对CPUE的间接影响,导致在探究环境因子与资源丰度关系时缺乏整体视角[16]。尽管路径分析或结构方程模型能够在一定程度解决这些问题[17−18],但这些方法对数据量和假设要求较高,通常适用于长期或大规模数据集[19]。相比之下,贝叶斯网络(Bayesian networks,BN)具有明显优势。它不仅能够结合定性和定量数据,灵活处理复杂的因果关系,还能揭示直接与间接的影响路径,同时对数据的要求相比其他方法较低[20]。近年来,贝叶斯网络在渔业研究中的应用逐渐增多[19–21],例如Araujo等[20]基于BN分析了影响鲑鱼早期存活率的潜在驱动因素,Malick等[21]则通过BN探索了气候变化等环境因素与鲑鱼补充量之间的联系。因此,贝叶斯网络成为了探索环境因子与渔业资源丰度之间联系的重要工具。
鉴于此,本文基于2014−2020年中国水产有限公司在马达加斯加西海岸捕虾拖网渔业数据,采用BN分析降水、径流、SST、SSHA和Chla环境因子与渔业主捕虾类CPUE之间的网络关系,旨在揭示这些因子及相互关系对虾类CPUE的影响路径、探索其影响机制;并探究BN在渔业资源研究环境因子与资源丰度分析中的适用性,为今后开展相关研究提供新的思路,为养护马达加斯加西海岸虾类资源、实现渔业可持续发展提供科学依据。
2. 材料与方法
2.1 数据来源
2.1.1 渔业数据
渔业数据来源于中国水产有限公司马达加斯加代表处2014−2020年统计的生产渔捞日志,渔船均采用臂架有翼单囊拖网作业方式,单船同时拖曳4顶结构相同的网具,网具(4片式)主尺度为38.0 m×32.5 m(17.0 m),网囊网目长度(2a)50 mm,并遵循当地渔业法律规定安装各类兼捕减少装置。限于当地渔业管理规定,渔业数据仅限于3−11月(12月至翌年2月为禁渔期),渔船作业范围为11°~21°S,43°~48°E,如图1所示。渔捞日志数据共计
20233 条,包含渔船放网和起网时间、渔船作业位置、拖曳时间和主要渔获种类渔获量等。 图 1 2014–2020年马达加斯加西海岸捕虾拖网渔场及CPUE分布Figure 1. Spatial distribution of fishing ground and CPUEs of shrimp trawl fisheries in west coast of Madagascar from 2014 to 2020
图 1 2014–2020年马达加斯加西海岸捕虾拖网渔场及CPUE分布Figure 1. Spatial distribution of fishing ground and CPUEs of shrimp trawl fisheries in west coast of Madagascar from 2014 to 2020使用CPUE作为指示虾类资源丰度指标。由于各船使用了相同规格的拖网渔具,且渔船作业习惯相近,因此使用一个渔区单元(0.5°×0.5°)内单位作业时长的渔获量作为渔业CPUE,即:
$$ {\mathrm{CPU{E}}}_{y,m,i}=\frac{\displaystyle\sum {C}_{y,m,i}}{\displaystyle\sum {E}_{y,m,i}}, $$ (1) 式中:
$ \displaystyle\sum {C}_{y,m,i} $ 为第y年第m月i单元渔区的总渔获量,单位为kg;$ \displaystyle\sum {E}_{y,m,i} $ 为为第y年第m月i单元渔区内总作业时长,单位为h。2.1.2 环境数据
淡水补充(降水和径流)数据来自NCEP/NCAR(美国国家环境预测中心/国家大气研究中心) 的再分析数据集网站
https://psl.noaa.gov/ ,采集数据的空间范围为10°~22°S,42°~50°E,空间分辨率为2.5°×2.5°,时间分辨率为月;SST、SSHA和Chla数据来自美国国家海洋和大气管理局(NOAA)的全球遥感数据网站https://coastwatch.pfeg.noaa.gov/ ,数据空间范围为10°~22°S,42°~50°E,空间分辨率为0.5°×0.5°,时间分辨率为月;并基于单元渔区的划分对SST、SSHA、Chla和降水、径流5个环境数据进行标准化处理,利用克里金空间插值法[22]对降水和径流数据进行了空间分辨率0.5°×0.5°重采样,其根据已知的大尺度降水和径流数据点赋权重预测小尺度未知点的值,得到最优化估计(表1)。表 1 环境变量的描述性统计Table 1. Descriptive statistics of environment variables环境变量 均值 标准差 最大值 最小值 降水/(mm∙d−1) 1.18 2.44 15.03 0 径流/(kg∙m−2) 0.40 0.98 5.31 0 海表面温度/℃ 27.54 1.63 30.82 23.72 海面高度异常/cm 0.06 0.05 0.34 −0.11 叶绿素a质量浓度/(mg∙L−1) 1.80 1.69 11.38 0.10 2.2 研究方法
2.2.1 贝叶斯网络介绍
贝叶斯网络(BN)也称为信念网络、因果网络或有向无环图模型,用以推断网络中变量之间的依赖性、确定变量之间的结构和关系[23−24]。BN由两个部分组成,即有向无环图(Directed Acyclic Graph, DAG)和条件概率表(Conditional Probability Tables, CPTs)。以一个简单的BN网络(图2)为例,其由3个节点代表变量(X1、X2和X3),由有向连接的边(X1→X2和X1→X3)表示它们之间的依赖关系,形成父级(X1)和子级(X2和X3)节点;而CPTs用来表示BN中所有相关联节点之间的可视化联合概率分布。
2.2.2 贝叶斯网络模型构建
贝叶斯网络以图形方式表示所选变量的联合概率分布,根据贝叶斯相关理论,节点X的联合概率可表示为
$$ f\left({x}_{1},\cdots ,{x}_{v}\right)=\prod_{i=1}^{v}f\left({x}_{i}|{\mathrm{\Pi }}_{{x}_{i}}\right) $$ (2) 式中:
$ f\left({x}_{1},\cdots ,{x}_{v}\right) $ 为联合概率;$ {\mathrm{\Pi }}_{{x}_{i}} $ 是$ {x}_{i} $ 父节点的先验概率(如果$ {x}_{i} $ 不存在父节点,那么$ {\mathrm{\Pi }}_{{x}_{i}} $ 表示为$ f\left({x}_{i}\right) $ );$ v $ 为节点数量。贝叶斯网络模型的构建可分为3步:(1)确定网络节点,即基于相关因果关系和专家经验确定对虾类CPUE产生影响的主要因素;(2)划分节点值,即对每个节点设置多个阈值区间表征不同的节点状态,通常需对数据进行离散化[25];(3)确定网络结构,常见构建方法包括由专家手动确定各边之间的位置和方向(因果贝叶斯网络),基于数据集采用机器学习算法创建网络结构(学习贝叶斯网络)以及结合上述两种方法的混合构建方法。
本研究采用混合方法构建贝叶斯网络,即首先根据相关海洋学知识和专家经验构建初始网络结构[22, 26],确定网络节点及其状态分布(图3和表2),然后使用基于分数的Hill-Climbing(HC)算法进行结构学习[27],采用最大似然估计进行参数学习估计局部分布函数的参数,得到各节点的条件概率,最终得到完整的贝叶斯网络模型。
 图 3 2014–2020年降水和海洋环境变量与马达加斯加西海岸主捕虾类的CPUE之间可能存在的关系Figure 3. Potential relationships between precipitation and ocean environmental variables with the CPUE of major shrimp species along the west coast of Madagascar from 2014 to 2020表 2 所有变量及其离散化区间Table 2. All variables and their discretization intervals
图 3 2014–2020年降水和海洋环境变量与马达加斯加西海岸主捕虾类的CPUE之间可能存在的关系Figure 3. Potential relationships between precipitation and ocean environmental variables with the CPUE of major shrimp species along the west coast of Madagascar from 2014 to 2020表 2 所有变量及其离散化区间Table 2. All variables and their discretization intervals降水/
(mm∙d−1)径流/
(kg∙m−2)海表面
温度/℃海表面高
度异常/cm叶绿素a质量浓度/
(mg∙L−1)印度白虾
WCPUE/
(kg∙h−1)独角新对虾
BCPUE/
(kg∙h−1)短沟对虾
TCPUE/
(kg∙h−1)x = 0 x = 0 x < 27 x < 0 x < 3 x < 10 x < 10 x < 5 x > 0 x > 0 x ≥ 27 x ≥ 0 x ≥ 3 x ≥ 10 x ≥ 10 x ≥ 5 使用R 4.4.0软件中的“bnlearn”包和GeNle软件实现网络结构的构建分析与可视化。
3. 结果
3.1 3种虾类CPUE和环境变量的月度变化
2014−2020年马达加斯加西海岸主捕虾类CPUE月平均变化如图4a–c所示。印度白虾月平均CPUE(WCPUE)整体呈现阶梯型下降趋势,其中3月月均CPUE值达到最大,随后逐月下降并趋于稳定;独角新对虾月平均CPUE(BCPUE)在3月至5月处于较高水平,随后逐月下降;短沟对虾月平均CPUE(TCPUE)整体呈现先上升后下降的趋势,在6月份达到峰值。
 图 4 2014–2020年马达加斯加西海岸3种虾类CPUE和环境变量的月平均变化Figure 4. Monthly average variations in CPUE of three shrimp species and environmental variables along the west coast of Madagascar from 2014 to 2020
图 4 2014–2020年马达加斯加西海岸3种虾类CPUE和环境变量的月平均变化Figure 4. Monthly average variations in CPUE of three shrimp species and environmental variables along the west coast of Madagascar from 2014 to 20202014−2020年马达加斯加西海岸海域的环境因子变化如图4d–h所示。降水和径流在3月份均达到最大值,随后呈减小趋势,其中降水在10−11月份出现回升;海表面温度整体呈先下降后上升趋势,变化范围从3月的30.8℃到7月的23.7℃;海面高度异常和叶绿素a质量浓度变化较小。
3.2 相关性分析
所有变量之间的相关性如图5所示。降水对印度白虾CPUE呈显著正相关(r = 0.44,p < 0.001),对独角新对虾也表现出显著正相关(r = 0.17,p < 0.001);径流对3种虾类都存在相关性,其中对印度白虾CPUE有高度正相关性(r = 0.66,p < 0.001)。SST对印度白虾和独角新对虾CPUE有显著正相关,对短沟对虾存在负相关,SSHA和Chla也对独角新对虾和短沟对虾CPUE有相关性。此外,降水与径流高度相关(r = 0.75,p < 0.001),SST与降水和径流也存在显著相关性,这些相关性表明马达加斯加西海岸海域的环境因子与淡水补充之间存在一定联系。
 图 5 所有变量之间的皮尔逊相关图***表示p < 0.001;**表示p < 0.01;*表示p < 0.05Figure 5. Pearson correlation plot between all variables*** indicates p < 0.001; **indicates p < 0.01; * indicates p < 0.05
图 5 所有变量之间的皮尔逊相关图***表示p < 0.001;**表示p < 0.01;*表示p < 0.05Figure 5. Pearson correlation plot between all variables*** indicates p < 0.001; **indicates p < 0.01; * indicates p < 0.053.3 贝叶斯网络分析
3.3.1 印度白虾
印度白虾的贝叶斯网络模型拟合结果(图6a和b)表明,预设的网络关系基本符合渔业实际情况,即印度白虾CPUE(WCPUE)受降水、径流、SST和SSHA的直接影响,而Chla对WCPUE无直接影响;降水除了对WCPUE的直接影响外,还会通过对径流、SST和SSHA产生影响进而对WCPUE产生间接影响。
 图 6 3种虾类的贝叶斯网络图(有向无环图及条件概率表,蓝色、红色箭头分别代表正显著和负显著关系)a. WCPUE(印度白虾)<10; b. WCPUE(印度白虾)≥10; c. BCPUE(独角新对虾)<10; d. BCPUE(独角新对虾)≥10; e. TCPUE(短沟对虾)<5; f. TCPUE(短沟对虾)≥5Figure 6. Bayesian Network of three shrimp species (DAGs and CPTs, blue and red arrows represent positive and negative significance relationship respectively)a. WCPUE(Fenneropenaeus indicus)<10; b. WCPUE(Fenneropenaeus indicus)≥10; c. BCPUE(Metapenaeus monoceros)<10; d. BCPUE(Metapenaeus monoceros)≥10; e. TCPUE(Penaeus semisulcatus)<5; f. TCPUE(Penaeus semisulcatus)≥5
图 6 3种虾类的贝叶斯网络图(有向无环图及条件概率表,蓝色、红色箭头分别代表正显著和负显著关系)a. WCPUE(印度白虾)<10; b. WCPUE(印度白虾)≥10; c. BCPUE(独角新对虾)<10; d. BCPUE(独角新对虾)≥10; e. TCPUE(短沟对虾)<5; f. TCPUE(短沟对虾)≥5Figure 6. Bayesian Network of three shrimp species (DAGs and CPTs, blue and red arrows represent positive and negative significance relationship respectively)a. WCPUE(Fenneropenaeus indicus)<10; b. WCPUE(Fenneropenaeus indicus)≥10; c. BCPUE(Metapenaeus monoceros)<10; d. BCPUE(Metapenaeus monoceros)≥10; e. TCPUE(Penaeus semisulcatus)<5; f. TCPUE(Penaeus semisulcatus)≥5基于不同WCPUE水平的条件概率表CPTs如图6a和b所示。对比低WCPUE和高WCPUE下降水发生的后验概率,即从86%提升至92%,表明降水对WCPUE具有显著的正向影响;径流是否存在的后验概率从低WCPUE下的45%增加至高WCPUE的79%,表明径流对WCPUE具有显著的正向影响;同理,从低WCPUE到高WCPUE,SSHA在正值区的后验概率下降(91%下降至85%),而SSHA负值区的后验概率上升(从9%提升至15%),表明SSHA对WCPUE具有显著的负向影响;SST≥27℃的后验概率从低WCPUE下的38%提升至高WCPUE下的86%,表明SST对WCPUE具有显著的正向影响。
3.3.2 独角新对虾
独角新对虾的贝叶斯网络模型拟合结果(图6c和d)表明径流、SST、SSHA和Chla对独角新对虾CPUE(BCPUE)产生直接影响,而降水对BCPUE无直接影响,但其可以通过径流、SST、SSHA和Chla对BCPUE产生间接影响。
基于不同BCPUE水平的条件概率表CPTs如图6c和d所示。SST≥27℃的后验概率从低BCPUE下的50%提升至高水平BCPUE下的60%,表明SST对BCPUE具有显著的正向影响,Chla≥3的后验概率从20%下降至12%,表明Chla对BCPUE具有显著的负向影响;类似的,径流的存在对BCPUE具有正向影响,SSHA对BCPUE负向影响。
3.3.3 短沟对虾
短沟对虾的贝叶斯网络模型拟合结果(图6e和f)表明径流、SST、SSHA和Chla对短沟对虾CPUE(TCPUE)产生直接影响,而降水对TCPUE无直接影响,但其可以通过径流、SST、SSHA和Chla对TCPUE产生间接影响。
基于不同TCPUE水平的条件概率表CPTs如图6e和f所示。对比低TCPUE和高TCPUE下径流存在的后验概率,即从62%下降至33%,表明径流存在对TCPUE具有显著负向影响;从低TCPUE到高TCPUE,SSHA处于正值区的后验概率下降(负值区的后验概率上升),表明SSHA对TCPUE具有显著负向影响;同理,SST和Chla对TCPUE具有显著负向影响。
4. 讨论
气候和海洋条件变化是虾类种群资源量变化的主要驱动因素,虾类资源丰度和种群分布对气候和海洋环境因子的响应机制尚不明确。本研究利用2014−2020年马达加斯加西海岸捕虾拖网数据,采用贝叶斯网络分析了降水、径流和SST等环境因子与3种虾类CPUE的影响路径和规律,探明了多环境因子影响对3种虾类CPUE在的潜在驱动因素,发现淡水补充是影响虾类CPUE的重要驱动因素,既可以直接影响虾类CPUE,也可以通过其他海洋环境因子对虾类CPUE产生间接影响。研究结果揭示了环境因子对渔业资源复杂而多层次的影响机制,这有助于更加全面地理解环境变化对马达加斯加西海岸沿海渔业的影响。
4.1 3种虾类CPUE的季节性变化
限于禁渔期等渔业管理规定,马达加斯加西海岸捕虾拖网仅在3−11月(12月至翌年2月为禁渔期)开展渔业生产。从2014−2020年间印度白虾CPUE随月份呈现显著下降的趋势,而独角新对虾和短沟对虾的CPUE呈先上升后下降趋势,这一差异表明马达加斯加西海岸3种主捕虾类资源的繁殖和补充在时间上并不同步。印度白虾的繁殖期主要为10−11月和3−4月[28],10−11月繁殖的幼虾在整个发育过程中,生长条件相对有利,自然和捕捞死亡率较低,这些幼虾在次年3−4月达成性成熟。然而,由于受环境条件改变的影响和渔业生产所造成的高捕捞死亡率,导致印度白虾CPUE从3月之后开始逐渐下降;独角新对虾的繁殖期在9−10月和5月[28],短沟对虾繁殖的高峰期在6月和12月[29],因此5月和6月分别为独角新对虾和短沟对虾资源量最为丰富的时期,出现较高CPUE。
4.2 环境因素及其相互关系对虾类CPUE的影响
近年来,关于淡水补充(降水和径流)产生的环境变化及其对渔业资源的影响已成为渔业研究中的热门话题[1, 7, 12, 13, 30]。降水是最重要的气候变量之一,其对水文循环的影响是最为直接的,一定程度上是水文循环的开始[31]。沿岸上升流、潮汐混合和陆地径流[32]是引起沿海区域养分富集的3个主要过程,其中径流不仅可以将陆地丰富的营养物质输送到海洋,还可以将幼鱼运送到近海,大流量的径流会降低仔幼鱼进入河口等水域的机会,是河口、沿岸渔业资源发生、补充和分布的重要驱动因素[1]。
此次研究发现,马达加斯加西海岸的淡水补充对捕虾拖网3种虾类CPUE具有影响,但降水和径流对不同虾类种群的影响机制可能存在差异。降水对印度白虾有显著的直接影响,但对独角新对虾和短沟对虾则主要通过径流等路径产生间接影响。径流对3种虾类均有显著的直接影响,并通过影响其它海洋条件对CPUE产生间接影响。径流对印度白虾和独角新对虾具有显著正向影响,但可能会抑制短沟对虾的资源丰度。马达加斯加西海岸每年的11月份至翌年的3月份为雨季,在这一季节贝齐布卡、齐里比希纳、曼古基和曼古鲁 4条河流(图1)大量淡水入海,栖息于近岸的虾类等种群资源量易受海洋条件变化影响,季节性降水及其引起的径流流量会把大量的营养物质、泥沙和虾类等带到海里,这些营养物质为虾类提供了丰富的饵料,使该水域幼鱼的补充量增加,对印度白虾和独角新对虾种群有着积极的影响[30, 33]。印度白虾和独角新对虾对盐度变化有更好的耐受性[34],而短沟对虾对盐度变化较为敏感[1],径流的增加导致沿岸地区盐度降低,可能会抑制短沟对虾的生长,进而引起短沟对虾CPUE的降低[35]。
SSHA反映涡流等中尺度特征,这些涡流通过多种机制影响生态系统的生产力。正负海面高度异常值不仅影响涡流的强度,还可能与温度变化、洋流和水质条件等多种因素相互作用[36]。Amores等[36]发现SSHA与虾类资源分布有显著的负相关性,并认为较高的SSHA增加了表层涡流活动进而增强底部附近水层的混合并将沉积物重新悬浮到水柱中,这一过程导致了渔场的浊度水平提高,水质条件的改变对虾类的摄食、活动等行为产生了多种负面影响,削弱了虾类的觅食能力,改变了虾类的行为习惯,降低了其被常规渔业方式的捕获率;SSHA通过改变循环模式,导致水温和洋流发生变化进而对虾类栖息分布造成影响。此外,由于混合的改变,SSHA的增大引起水柱分层增加会限制海水垂直混合,形成更为明显的温度梯度,驱使虾类向下移动以寻找适宜栖息空间。如布埃纳文图拉沿海的SSHA与西方白虾(Litopenaeus occidentalis)CPUE呈负相关[7]。本文研究也证实了这一影响机制,SSHA对3种虾类CPUE均产生负向影响。
此外,BN网络的分析结果表明,SSHA也受降水和径流影响,表明径流和降水通过SSHA对虾类分布产生间接影响。SSHA通常受风应力、热通量和淡水补充的影响[37],淡水补充通过产生正压信号迅速分散到世界海洋中[38],且淡水补充还可以通过进入海洋等温层,导致混合层变浅,对SSHA产生影响[39],这可能导致主捕虾类的栖息地发生改变,最终对捕虾拖网渔业的CPUE造成影响。
水温能直接或者间接影响虾类等水生变温动物的生活习性[40],是影响种群分布的是关键环境因子,也是渔场分析和预测的重要因素。虾类对温度变化非常敏感[41],水温降低时虾类会出现潜底等行为[42]。本文研究发现SST对印度白虾和独角新对虾的CPUE有显著的正向影响,这与马鞍列岛海域优势甲壳类CPUE对SST的响应规律一致[43]。然而,SST对短沟对虾CPUE有显著的负相关影响,表明不同虾类对温度的响应可能存在差异。
此外,SST还与降水、径流和SSHA存在相互作用,进而对CPUE产生一定影响。对于降水、径流和SST的关系,大量研究集中于降水与SST的相互作用。Wu[44]发现季节内降水扰动与降水引起的海表面温度变化的相关性随着总降雨量的减小而减小。Chen等[45]通过研究20世纪90年代末南海同期降水和SST数据,发现南海西南海域降水与SST呈显著正相关。本文研究发现降水与SST的正相关关系以及BN网络结果都表明马达加斯加西海岸降水和径流等通过SST对虾类CPUE产生间接影响。
Chla浓度是表征浮游植物丰富程度的一个重要指标,广泛用于描述生态系统的生产力与食物链之间的关系[46]。尽管在一定范围内,Chla增加可以提升饵料生物丰度进而吸引鱼类等群体聚集[47],提高渔业CPUE。但也有研究发现莫桑比克海峡的独角新对虾和短沟对虾CPUE与Chla整体呈负相关[48],这与本研究结果基本一致。与普遍机制存在差异原因可能与马达加斯加西海岸海域特征有关,这一区域季节性气候变化以及河流径流可能会导致水体中的浮游植物过度繁殖[49],增加Chla,进而引发局部缺氧或低氧环境,不利于虾类的生长和分布,特别是当水体的生物生产力超出其生态平衡点会导致饵料资源结构的改变或食物竞争加剧,从而抑制虾类CPUE。
4.3 贝叶斯网络的适用性和局限性
本研究基于贝叶斯网络成功构建了降水等环境因素对马达加斯加西海岸捕虾拖网虾类CPUE的影响路径,表明通过BN揭示影响因素的相互关系,从而更全面地理解其对响应变量的潜在影响机制是可行的。研究结果证实了环境因素对虾类种群丰度不仅存在显著的直接影响,还可以通过多种路径产生间接影响。与传统统计模型相比,BN能够揭示因子之间复杂的相互关系,突破了仅关注于独立变量及交互作用的局限性,是对传统方法的有效补充。因此,BN在渔业资源管理中的应用可为进一步探索环境因素对渔业资源变动机制提供新的思路和参考。
尽管贝叶斯网络是一个强大且灵活的工具,能够有效阐明环境因子对渔业资源量影响的直接和间接途径,但也存在一定的局限性。首先,本研究未考虑环境因子的时间滞后效应,这意味着未能充分捕捉这些因素对渔业资源量变化的延迟影响。其次,贝叶斯网络的推理过程往往依赖于先验知识的选择和专家判断,因此预设的网络结构可能带有一定的主观性,从而影响模拟结果。为了逐步消除这些局限性,在未来的研究中我们计划引入滞后效应分析,例如采用贝叶斯网络的扩展——动态贝叶斯网络,以期更好揭示渔业资源变化对环境的延迟响应。此外,我们还将进一步增强数据驱动的模型构建,减少对专家判断的依赖,具体来说,我们将通过结构学习算法自动学习变量之间的关系,并结合先验知识和专家判断,逐步减少主观判断对模型结构的影响。
-

图 1 2014–2020年马达加斯加西海岸捕虾拖网渔场及CPUE分布
Fig. 1 Spatial distribution of fishing ground and CPUEs of shrimp trawl fisheries in west coast of Madagascar from 2014 to 2020

图 3 2014–2020年降水和海洋环境变量与马达加斯加西海岸主捕虾类的CPUE之间可能存在的关系
Fig. 3 Potential relationships between precipitation and ocean environmental variables with the CPUE of major shrimp species along the west coast of Madagascar from 2014 to 2020

图 4 2014–2020年马达加斯加西海岸3种虾类CPUE和环境变量的月平均变化
Fig. 4 Monthly average variations in CPUE of three shrimp species and environmental variables along the west coast of Madagascar from 2014 to 2020

图 5 所有变量之间的皮尔逊相关图
***表示p < 0.001;**表示p < 0.01;*表示p < 0.05
Fig. 5 Pearson correlation plot between all variables
*** indicates p < 0.001; **indicates p < 0.01; * indicates p < 0.05
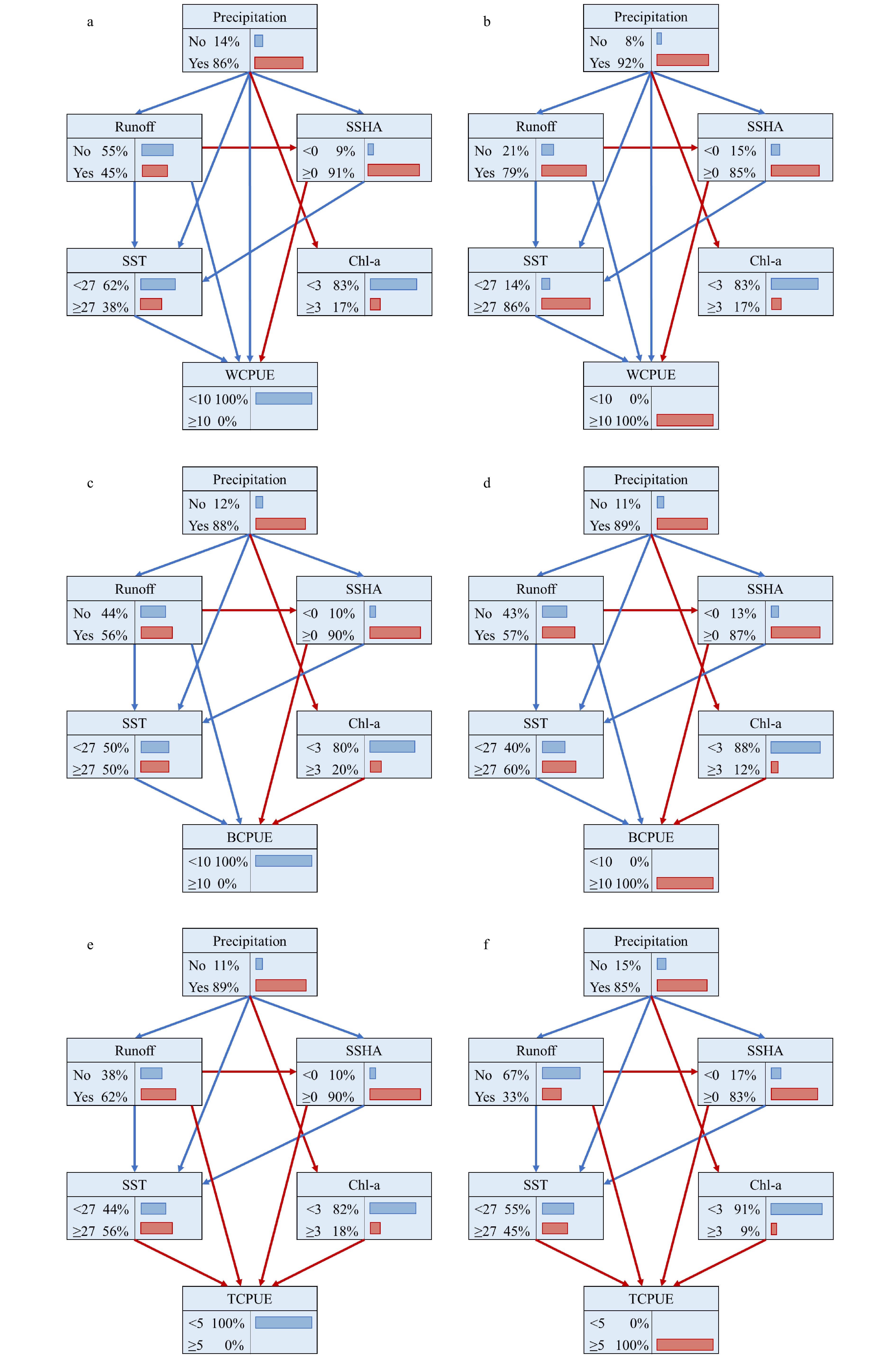
图 6 3种虾类的贝叶斯网络图(有向无环图及条件概率表,蓝色、红色箭头分别代表正显著和负显著关系)
a. WCPUE(印度白虾)<10; b. WCPUE(印度白虾)≥10; c. BCPUE(独角新对虾)<10; d. BCPUE(独角新对虾)≥10; e. TCPUE(短沟对虾)<5; f. TCPUE(短沟对虾)≥5
Fig. 6 Bayesian Network of three shrimp species (DAGs and CPTs, blue and red arrows represent positive and negative significance relationship respectively)
a. WCPUE(Fenneropenaeus indicus)<10; b. WCPUE(Fenneropenaeus indicus)≥10; c. BCPUE(Metapenaeus monoceros)<10; d. BCPUE(Metapenaeus monoceros)≥10; e. TCPUE(Penaeus semisulcatus)<5; f. TCPUE(Penaeus semisulcatus)≥5
表 1 环境变量的描述性统计
Tab. 1 Descriptive statistics of environment variables
环境变量 均值 标准差 最大值 最小值 降水/(mm∙d−1) 1.18 2.44 15.03 0 径流/(kg∙m−2) 0.40 0.98 5.31 0 海表面温度/℃ 27.54 1.63 30.82 23.72 海面高度异常/cm 0.06 0.05 0.34 −0.11 叶绿素a质量浓度/(mg∙L−1) 1.80 1.69 11.38 0.10  下载: 导出CSV
下载: 导出CSV
表 2 所有变量及其离散化区间
Tab. 2 All variables and their discretization intervals
降水/
(mm∙d−1)径流/
(kg∙m−2)海表面
温度/℃海表面高
度异常/cm叶绿素a质量浓度/
(mg∙L−1)印度白虾
WCPUE/
(kg∙h−1)独角新对虾
BCPUE/
(kg∙h−1)短沟对虾
TCPUE/
(kg∙h−1)x = 0 x = 0 x < 27 x < 0 x < 3 x < 10 x < 10 x < 5 x > 0 x > 0 x ≥ 27 x ≥ 0 x ≥ 3 x ≥ 10 x ≥ 10 x ≥ 5
下载: 导出CSV
-
[1] Caverivière A, Chaboud C, Rafalimanana T, et al. Les Crevettes Côtières de Madagascar: Biologie, Exploitation, Gestion[M]. Marseille: IRD Éditions, 2017. [2] Fourmanoir P. Nouvelles expériences de pêche du Gabriel-II effectuées du 24 juillet[J]. Bulletin de Madagascar, 1952(52): 17−21. [3] Farfante I P, Kensley B. Penaeoid and Sergestoid Shrimps and Prawns of the World[M]. Paris: Mémoires Du Museum National D'histoire Naturelle, 1997. [4] De Young C. Review of the State of World Marine Capture Fisheries Management: Indian Ocean / Edited by Cassandra de Young[M]. Rome: Food and Agriculture Organization of the United Nations, 2006. [5] Manach F L, Gough C, Harris A, et al. Unreported fishing, hungry people and political turmoil: the recipe for a food security crisis in Madagascar?[J]. Marine Policy, 2012, 36(1): 218−225 [6] Aeinjamshid K, Fazelpoor K, Rabbaniha M, et al. Identification of shrimp potential fishing zones in the northern part of the Persian Gulf using the Fuzzy Overlay model for integrating satellite and ground-based data[J]. Iranian Journal of Fisheries Sciences, 2023, 22(4): 907−924. [7] Díaz-Ochoa J A, Quiñones R A. Relationship of precipitation, freshwater input, and sea level height with the abundance of the white shrimp (Litopenaeus occidentalis; Street, 1871) off Buenaventura, eastern tropical Pacific[J]. Fisheries Research, 2008, 92(2−3): 148−161. doi: 10.1016/j.fishres.2008.01.002 [8] Grimes C B. Fishery production and the Mississippi River discharge[J]. Fisheries, 2001, 26(8): 17−26. doi: 10.1577/1548-8446(2001)026<0017:FPATMR>2.0.CO;2 [9] Erzini K. Trends in NE Atlantic landings (southern Portugal): identifying the relative importance of fisheries and environmental variables[J]. Fisheries Oceanography, 2005, 14(3): 195−209. doi: 10.1111/j.1365-2419.2005.00332.x [10] Doney S C. The growing human footprint on coastal and open-ocean biogeochemistry[J]. Science, 2010, 328: 1512−1516. doi: 10.1126/science.1185198 [11] Najjar R G, Pyke C R, Adams M B, et al. Potential climate-change impacts on the Chesapeake Bay[J]. Estuarine, Coastal and Shelf Science, 2010, 86(1): 1−20. doi: 10.1016/j.ecss.2009.09.026 [12] Gracia A. Relationship between environmental factors and white shrimp abundance in the southwestern Gulf of Mexico[J]. Anales del Instituto Ciencias del Mar y Limnología, 1989, 16(1): 171−182. [13] Evans C, Opnai L J, Kare B D. Fishery ecology and oceanography of the prawn Penaeus merguiensis (de Man) in the Gulf of Papua: estimation of maximum sustainable yield and modelling of yield, effort and rainfall[J]. Marine and Freshwater Research, 1997, 48(3): 219−228. doi: 10.1071/MF94222 [14] Tucker S, Thiess M S, Morris J F T, et al. Coastal distribution and consequent factors influencing production of endangered Snake River sockeye salmon[J]. Transactions of the American Fisheries Society, 2015, 144(1): 107−123. doi: 10.1080/00028487.2014.968292 [15] 唐浩, 许柳雄, 陈新军, 等. 基于GAM模型研究时空及环境因子对中西太平洋鲣鱼渔场的影响[J]. 海洋环境科学, 2013, 32(4): 518−522.Tang Hao, Xu Liuxiong, Chen Xinjun, et al. Effects of spatiotemporal and environmental factors on the fishing ground of skipjack tuna (Katsuwonus pelamis) in the Western and Central Pacific Ocean based on generalized additive model[J]. Marine Environmental Science, 2013, 32(4): 518−522. [16] Gu Yixin, Wang Lifei, Lyu S L, et al. Assessing potential driving factors of the ecosystem service value of mariculture shellfish in China using a structural equation modeling approach[J]. Frontiers in Marine Science, 2023, 10: 1232211. doi: 10.3389/fmars.2023.1232211 [17] 廖祖军, 王学锋, 周艳波, 等. 基于结构方程模型分析海洋环境因子对南海鸢乌贼的影响[J]. 南方水产科学, 2024, 20(2): 11−18.Liao Zujun, Wang Xuefeng, Zhou Yanbo, et al. Analysis of effects of environmental factors on Sthenoteuthis oualaniensis based on structural equation model[J]. South China Fisheries Science, 2024, 20(2): 11−18. [18] 张颖, 王旭红, 冯子豪, 等. 基于SEM模型的热环境时空特征及影响因素分析: 以西安都市圈为例[J]. 环境科学, 2024, 45(6): 3734−3745.Zhang Ying, Wang Xuhong, Feng Zihao, et al. Spatiotemporal pattern and influencing factors of thermal environment based on SEM model: a case study in Xi'an metropolitan area[J]. Environmental Science, 2024, 45(6): 3734−3745. [19] Hertz E, Trudel M, Tucker S, et al. Influences of ocean conditions and feeding ecology on the survival of juvenile Chinook Salmon (Oncorhynchus tshawytscha)[J]. Fisheries oceanography, 2016, 25(4): 407−419. doi: 10.1111/fog.12161 [20] Araujo H A, Holt C, Curtis J M, et al. Building an ecosystem model using mismatched and fragmented data: a probabilistic network of early marine survival for coho salmon Oncorhynchus kisutch in the Strait of Georgia[J]. Progress in Oceanography, 2013, 115: 41−52. doi: 10.1016/j.pocean.2013.05.022 [21] Malick M J, Cox S P, Peterman R M, et al. Accounting for multiple pathways in the connections among climate variability, ocean processes, and coho salmon recruitment in the Northern California Current[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2015, 72(10): 1552−1564. doi: 10.1139/cjfas-2014-0509 [22] 马有成, 黄斌, 张健, 等. 马达加斯加西海岸3种虾类CPUE时空分布及其与环境因子关系[J]. 海洋渔业, 2024, 46(5): 540−550.Ma Youcheng, Huang Bin, Zhang Jian, et al. Temporal and spatial distribution of CPUE in three species of shrimps and its relationship with environmental factors along the west coast of Madagascar[J]. Marine Fisheries, 2024, 46(5): 540−550. [23] Nagarajan R, Scutari M, Lèbre S. Bayesian Networks in R[M]. New York: Springer, 2013, 122: 125-127. [24] Pearl J. Bayesian networks A model of self-activated memory for evidential reasoning[C]//Proceedings of the 7th Conference of the Cognitive Science Society. Irvine: University of California, 1985: 15−17. [25] Uusitalo L. Advantages and challenges of Bayesian networks in environmental modelling[J]. Ecological modelling, 2007, 203(3/4): 312−318. [26] 张省宇. 马达加斯加西海岸海洋环境因子与主要虾类CPUE的关系[D]. 上海: 上海海洋大学, 2015.Zhang Shengyu. The relation between marine environment factor and the CPUE of main shrimp and prawn in inshore west of Madagascar[D]. Shang Hai: Shanghai Ocean University, 2015. [27] Scutari M. Bayesian network constraint-based structure learning algorithms: parallel and optimized implementations inthe bnlearn R Package[J]. Journal of Statistical Software, 2017, 77(2): 1−20. [28] Le Reste L, Marcille J. Biologie de la crevette Penaeus indicus H. Milne Edwards à Madagascar: croissance, recrutement, migrations reproduction, mortalité: contribution à l'étude d'une baie eutrophique tropicale[J]. Cahiers ORSTOM. Série Océanographie, 1976, 14(2): 109−127. [29] Crocos P J, Van der Velde T D. Seasonal, spatial and interannual variability in the reproductive dynamics of the grooved tiger prawn Penaeus semisulcatus in Albatross Bay, Gulf of Carpentaria, Australia: the concept of effective spawning[J]. Marine Biology, 1995, 122(4): 557−570. doi: 10.1007/BF00350678 [30] Whitfield A. A characterization of southern African estuarine systems[J]. Southern African Journal of Aquatic Sciences, 1992, 24(1/2): 86−98. [31] Chang Jian, Wei Jie, Wang Yimin, et al. Precipitation and runoff variations in the Yellow River Basin of China[J]. Journal of Hydroinformatics, 2017, 19(1): 138−155. doi: 10.2166/hydro.2016.047 [32] Caddy J F, Bakun A. A tentative classification of coastal marine ecosystems based on dominant processes of nutrient supply[J]. Ocean & Coastal Management, 1994, 23(3): 201−211. [33] Cowley P, Whitfield A, Bell K. The surf zone ichthyoplankton adjacent to an intermittently open estuary, with evidence of recruitment during marine overwash events[J]. Estuarine, Coastal and Shelf Science, 2001, 52(3): 339−348. doi: 10.1006/ecss.2000.0710 [34] Vanza J G, Solanki H G, Borichangar R V, et al. Effect of different salinities levels on growth and survival of Indian white shrimp (Fenneropenaeus indicus)[J]. International Journal of Current Microbiology and Appied Sciences, 2018, 7(7): 2295−2304. doi: 10.20546/ijcmas.2018.707.268 [35] Niamaimandi N, Arshad A B, Daud S K, et al. Population dynamic of green tiger prawn, Penaeus semisulcatus (De Haan) in Bushehr coastal waters, Persian Gulf[J]. Fisheries Research, 2007, 86(2/3): 105−112. [36] Amores A, Rueda L, Monserrat S, et al. Influence of the hydrodynamic conditions on the accessibility of Aristeus antennatus and other demersal species to the deep water trawl fishery off the Balearic Islands (western Mediterranean)[J]. Journal of Marine Systems, 2014, 138: 203−210. doi: 10.1016/j.jmarsys.2013.11.014 [37] Zhang Xiaolin, Takashi M. Climate mode impacts on the sea surface height anomaly in the Indian Ocean[J]. ESS Open Authorea Preprints, 2022. [38] Huang Ruixin, Jin Xiangze. Sea surface elevation and bottom pressure anomalies due to thermohaline forcing. Part I: Isolated perturbations[J]. Journal of Physical Oceanography, 2002, 32(7): 2131−2150. doi: 10.1175/1520-0485(2002)032<2131:SSEABP>2.0.CO;2 [39] Zhang Xiaolin, Clarke A J. Observations of interannual equatorial freshwater jets in the western Pacific[J]. Journal of Physical Oceanography, 2015, 45(11): 2848−2865. doi: 10.1175/JPO-D-14-0245.1 [40] Cota-Durán A, Petatán-Ramírez D, Ojeda-Ruiz M Á, et al. Potential impacts of climate change on shrimps distribution of commercial importance in the Gulf of California[J]. Applied Sciences, 2021, 11(12): 5506. doi: 10.3390/app11125506 [41] El-Damhogy K A E, Heneash A M M, Ahmed E. Impact of water temperature and salinity on the distribution and abundance of shrimp (Crustacean: Decapoda) at Lake Burullus, Egypt[J]. International Journal of Ecotoxicology and Ecobiology, 2016, 2(1): 1−7. [42] Rao G S, Radhakrishnan E, Josileen J. Handbook of Marine Prawns of India[M]. Kochi: ICAR-Central Marine Fisheries Research Institute, 2013. [43] 赵静, 梁金玲, 周曦杰, 等. 基于GAM模型的马鞍列岛海域优势甲壳类与环境因子的关系研究[J]. 南方水产科学, 2017, 13(3): 26−35.Zhao Jing, Liang Jinling, Zhou Xijie, et al. Research on relationship between dominant crustacean species and environmental factors based on GAM[J]. South China Fisheries Science 2017, 13(3): 26−35. [44] Wu Renguang. Summer precipitation–SST relationship on different time scales in the northern tropical Indian Ocean and western Pacific[J]. Climate Dynamics, 2019, 52(9/10): 5911−5926. [45] Chen Jiepeng, Wang Xin, Zhou Wen, et al. Interdecadal change in the summer SST-precipitation relationship around the late 1990s over the South China Sea[J]. Climate Dynamics, 2018, 51(5/6): 2229−2246. [46] Druon J N, Chassot E, Murua H, et al. Preferred feeding habitat of skipjack tuna in the eastern central Atlantic and western Indian Oceans: relations with carrying capacity and vulnerability to purse seine fishing[C]. Seychelles: IOTC Proceedings. IOTC, 2016: 18−31. [47] Syah A, Ni’am A, Jatisworo D. Potential fishing grounds of Skipjack tuna (Katsuwonus pelamis) in western water of Sumatera using remotely sensed data and maximum entropy model[J]. Bristol: IOP Conference Series: Earth and Environmental Science, 2023, 1251(1): 012066. doi: 10.1088/1755-1315/1251/1/012066 [48] 王建城. 莫桑比克海峡捕虾拖网渔场时空分布及虾类资源丰度与环境因子的关系[D]. 上海: 上海海洋大学, 2023.Wang Jiancheng. Spatial and temporal distribution of shrimp trawl fishing grounds in the Mozambique Channel and the relationship between shrimpre source abundance and environmental factor[D]. Shang Hai: Shanghai Ocean University, 2023. [49] Shafeeque M, Shah P, Platt T, et al. Effect of precipitation on chlorophyll-a in an upwelling dominated region along the west coast of India[J]. Journal of Coastal Research, 2019, 86(sp1): 218−224. doi: 10.2112/SI86-032.1 -
 下载:
下载:
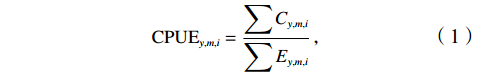
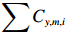
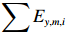

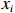
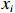
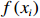
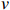
计量
- 文章访问数: 70
- HTML全文浏览量: 37
- PDF下载量: 7
- 被引次数: 0
