

Characteristics and their influencing factors of marine nematode communities in mangrove wetlands of Sanya City, Hainan Province
-
摘要: 本研究于2020年11月在海南亚龙湾青梅港红树林自然保护区和三亚河红树林自然保护区采集沉积物样品,开展了海洋线虫的群落特征及影响因子研究。研究结果显示,青梅港红树林海洋线虫平均丰度为(80.4 ± 40.3) ind./cm2,三亚河红树林的为(96.7 ± 55.6) ind./cm2,两处红树林湿地海洋线虫平均丰度为(88.5 ± 47.1) ind./cm2,海洋线虫丰度在两红树林湿地间存在显著差异。共鉴定海洋线虫77种,隶属于56属,21科7目。青梅港红树林的优势属为新色矛线虫属(Neochromadora)、微口线虫属(Terschellingia)、拟单宫线虫属(Paramonohystera)、吞咽线虫属(Daptonema)、链环线虫属(Desmodora),以刮食性线虫为优势摄食类型;三亚河红树林的优势属为微口线虫属(Terschellingia)、萨巴线虫属(Sabatieria)、游咽线虫属(Eleutherolaimus)、拟齿线虫属(Parodontophora)和折咽线虫属(Ptycholaimellus),以选择性沉积食性线虫为主;两红树林湿地共同的优势属为微口线虫属。海洋线虫群落结构在两红树林均具空间差异,海洋线虫群落在两红树林之间分化,但差异并不显著。沉积物有机质含量和脱镁叶绿酸含量是影响三亚河红树林海洋线虫丰度和群落结构的重要因素;红树植被类型的差异可能在一定程度上对青梅港红树林海洋线虫的丰度及群落结构产生影响;沉积物粒度特征、海水pH值和盐度等的差异可解释两处红树林选定站点海洋线虫群落的分化。此外,青梅港红树林存在污水工程、港口建设和旅游业的发展,三亚河红树林存在人为砍伐、筑堤、盐田开辟、养虾池废水排放,两地存在的不同污染也可能是导致红树林海洋线虫群落分化的原因。Abstract: In November 2020, sediment samples were collected from the Yalong Bay Qingmei Port mangrove nature reserve and the Sanya River mangrove nature reserve in Hainan Province. The present study focused on the community characteristics of marine nematodes and their influencing factors in these two mangroves. The results of the present study showed that the average abundance of marine nematodes was (80.4 ± 40.3) ind./cm2 in the Qingmei Port mangrove, while it was (96.7 ± 55.6) ind./cm2 in the Sanya River mangrove. In this study, the average abundance of marine nematodes in the two study mangrove wetlands was (88.5 ± 47.1) ind./cm2. There were significant differences in the abundance of marine nematodes between the two mangrove wetlands. A total of 77 species of marine nematodes were identified, belonging to 56 genera, 21 families and 7 orders. The dominant genera in the Qingmei Port mangrove were Neochromadora, Terschellingia, Paramonohystera, Daptonema and Desmodora, and epigrowth feeders were the dominant feeding type. The dominant genera in the Sanya River mangrove were Terschellingia, Sabatieria, Eleutherolaimus, Parodontophora and Ptycholaimellus, and the selective deposit feeders dominated. Terschellingia dominated in both mangroves. There were spatial differences in the community structure of marine nematodes in study mangroves. Although the marine nematode communities differentiated between the two mangroves, the difference was not significant. Organic matter content and phaeophorbide content in sediments were important factors affecting the abundance and community structure of marine nematodes in Sanya River mangrove. The differences in mangrove vegetation types may affect the abundance and community structure of marine nematodes in Qingmei Port mangrove to some extent. The differences of sediment granularity characteristics, seawater pH and salinity could explain the differentiation of marine nematode communities at selected sites of the two mangrove. In addition, there are sewage project, port construction and tourism development in Qingmei Port mangrove. And there are felling, dike construction, salt pan development and wastewater discharge from shrimp ponds in Sanya River mangrove. The different pollution status in these two mangroves may also be the reason for the differences in the abundance and community structure of marine nematodes in this study.
-
Key words:
- mangrove /
- marine nematode /
- species composition /
- feeding structure /
- Sanya
-

图 2 研究站点环境因子主成分分析(PCA)
Fig. 2 Principal component analysis (PCA) result of environmental factors in the study sites
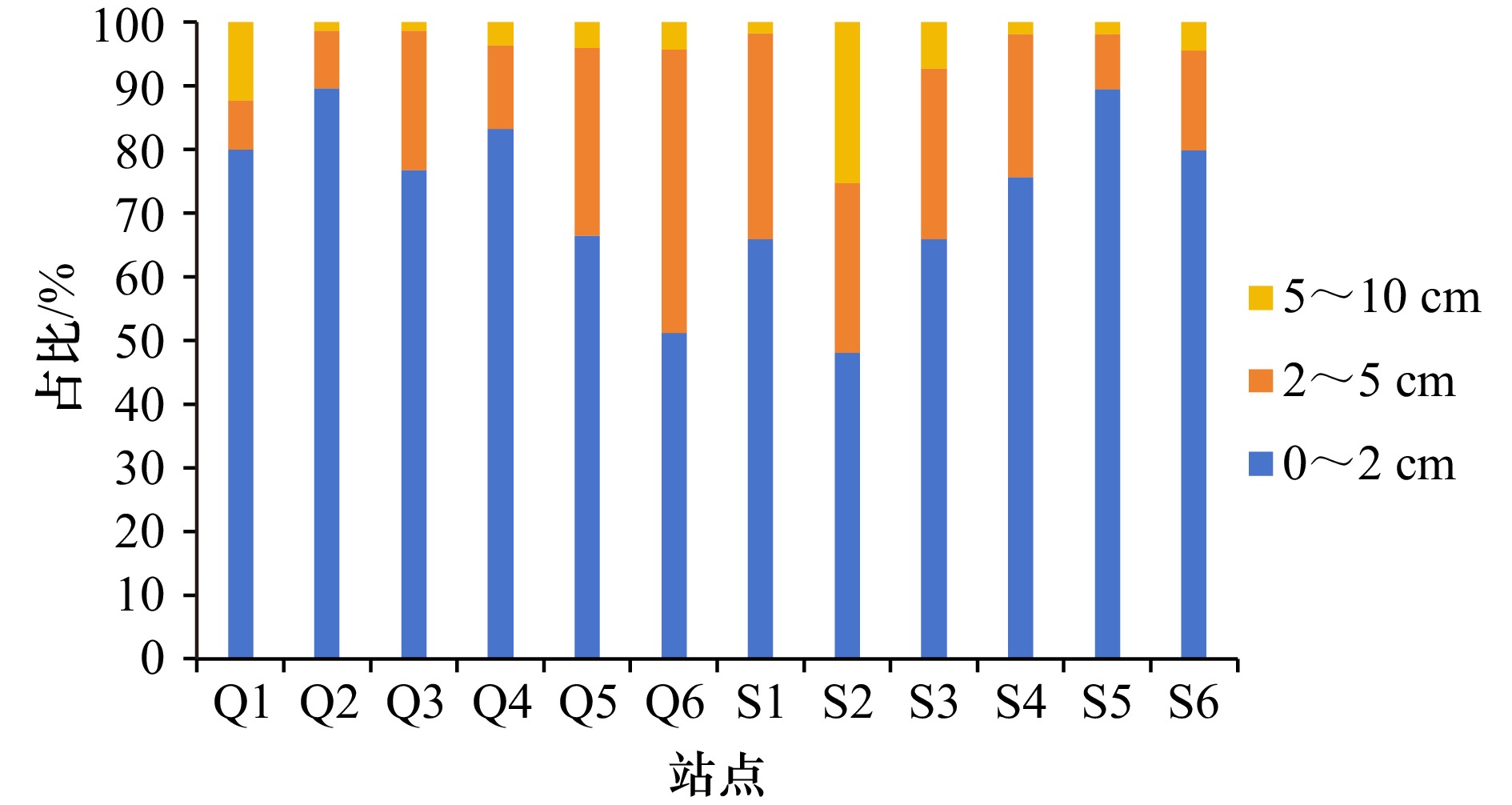
图 3 两个红树林湿地海洋线虫的垂直分布
Fig. 3 Vertical distribution of marine nematodes in the two mangrove wetlands

图 4 两个红树林湿地海洋线虫群落的非度量多维度分析
Fig. 4 Metric multidimensional scaling plot of marine nematode communities in the two mangrove wetlands
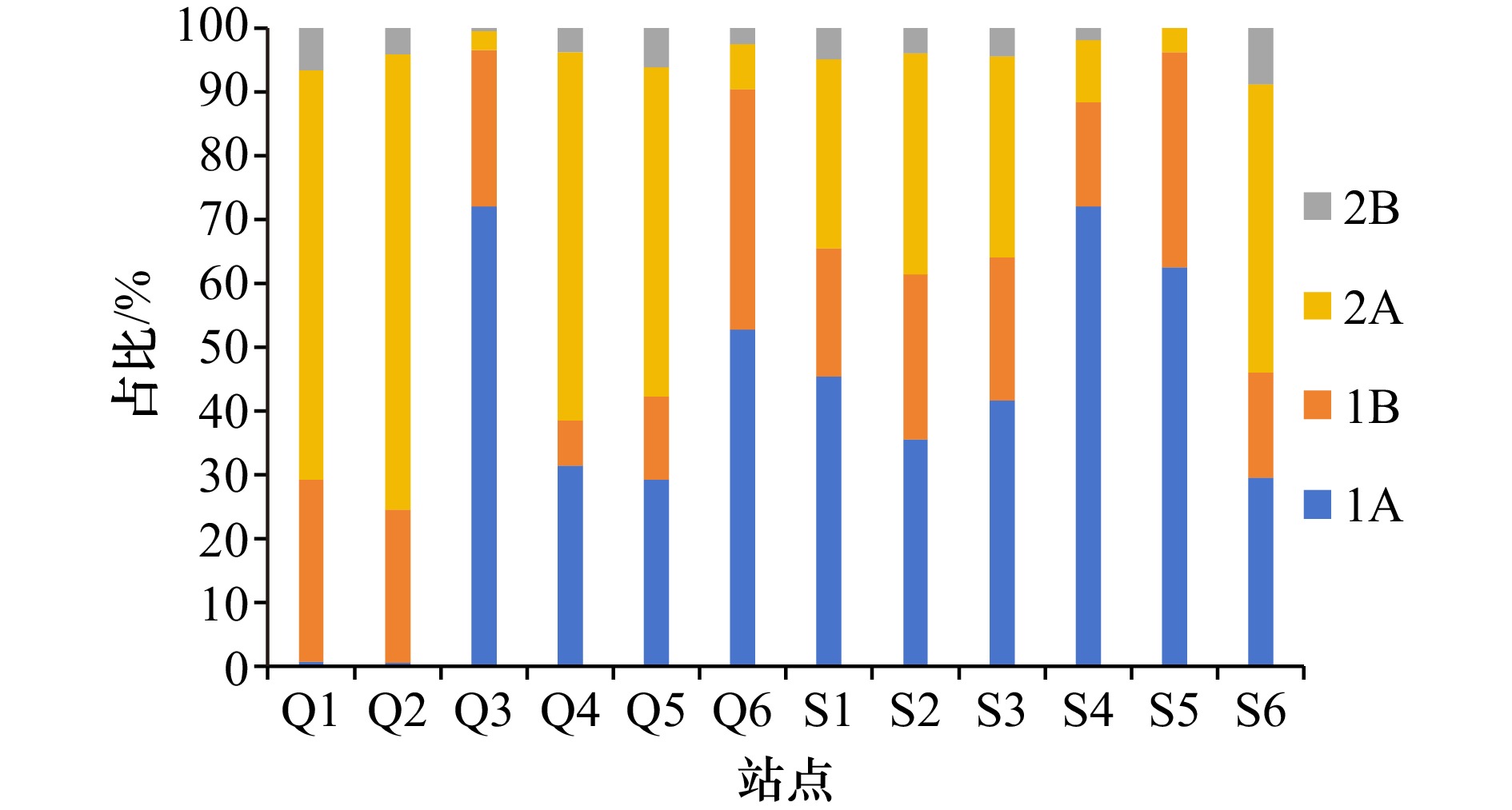
图 5 两个红树林湿地海洋线虫的摄食类型组成
Fig. 5 Proportion of feeding types of marine nematodes in the two mangrove wetlands
表 1 研究站点环境因子
Tab. 1 Environmental factors of study sites
研究区域 采样点 pH DO/
(mg∙L−1)T/℃ S σi Md/μm OM/% Chl-a/
(μg∙g−1)Pheo-a/
(μg∙g−1)TY/% Tannin/
(mg∙g−1)青梅港 Q1 7.87 5.37 28.38 3.13 1.30 332.48 2.03 0.29 0.69 1.44 20.41 Q2 1.32 358.79 0.80 0.99 1.30 0.66 33.70 Q3 7.91 5.69 28.80 3.88 1.32 426.92 8.38 0.17 0.43 1.63 20.22 Q4 1.36 360.33 1.64 0.32 0.99 2.01 19.70 Q5 7.68 5.27 30.04 3.72 2.93 41.49 7.13 0.41 1.75 57.33 21.70 Q6 1.36 392.35 1.33 0.43 1.45 2.00 20.09 平均 7.82 5.44 29.07 3.58 1.60 318.73 3.55 0.43 1.10 10.84 22.64 三亚河 S1 8.21 6.14 26.91 14.12 2.34 119.96 5.86 0.50 1.73 27.08 41.45 S2 1.12 153.43 3.99 0.79 1.50 23.48 27.70 S3 1.61 115.76 3.18 0.25 0.76 19.87 20.74 S4 8.12 5.64 26.81 13.05 2.42 118.27 2.73 0.63 1.98 27.54 23.96 S5 2.83 100.71 9.52 0.52 2.75 33.42 24.22 S6 3.08 64.70 9.05 0.86 3.77 49.30 19.06 平均 8.17 5.89 26.86 13.59 2.23 112.14 5.72 0.59 2.08 30.11 26.19 注:pH:酸碱度;DO:溶解氧含量;T:温度;S:盐度;σi:分选系数;Md:中值粒径;OM:有机质含量;Chl-a:叶绿素含量;Pheo-a:脱镁叶绿酸含量;TY:粉砂黏土含量;Tannin:单宁含量。  下载: 导出CSV
下载: 导出CSV
表 2 两个红树林湿地的海洋线虫丰度
Tab. 2 Abundance of marine nematodes in the two mangrove wetlands
研究站点 丰度/
(ind.∙cm−2)研究站点 丰度/
(ind.∙cm−2)Q1 61.0 ± 23.2 S1 144.4 ± 39.3 Q2 125.5 ± 88.1 S2 122.4 ± 92.3 Q3 74.8 ± 52.2 S3 139.4 ± 73.5 Q4 134.5 ± 65.4 S4 122.1 ± 72.3 Q5 42.9 ± 23.8 S5 23.0 ± 23.3 Q6 43.6 ± 43.3 S6 28.7 ± 23.9 平均 80.4 ± 40.3 平均 96.7 ± 55.6
下载: 导出CSV
表 3 两个红树林湿地的海洋线虫优势属(优势度≥5%)
Tab. 3 Dominant genera of marine nematodes in the two mangrove wetlands (dominance ≥5%)
青梅港 属名 Q1 Q2 Q3 Q4 Q5 Q6 总 Basiria 22.42% 6.90% Daptonema 5.76% 20.33% 28.93% 8.72% Desmodora 5.02% 30.30% 8.52% Haliplectus 9.09% Linhomoeus 22.55% Neochromadora 43.89% 30.67% 23.94% 15.95% 22.44% Paracanthonchus 26.36% Paracyatholaimus 10.48% 5.76% Paramonohystera 22.27% 13.57% 8.85% Sabatieria 7.55% Terschellingia 71.74% 18.71% 52.20% 19.93% Trissonchulus 5.21% Viscosia 6.55% 三亚河 属名 S1 S2 S3 S4 S5 S6 总 Chromadorella 5.49% Daptonema 7.40% 5.05% 6.25% 10.99% Eleutherolaimus 6.40% 13.01% 8.87% 7.05% Haliplectus 6.25% Metadesmolaimus 8.40% 5.00% Oxystomina 6.25% 7.69% Parodontophora 6.08% 8.94% 7.15% 6.33% Ptycholaimellus 10.69% 7.31% 5.45% Sabatieria 10.23% 13.82% 11.57% 14.33% 11.41% Southerniella 15.00% Terschellingia 42.00% 33.06% 38.67% 72.21% 50.00% 13.19% 44.71% Theristus 7.50% Trissonchulus 26.37%
下载: 导出CSV
表 4 两个红树林湿地的海洋线虫优势种(优势度≥5%)
Tab. 4 Dominant species of marine nematodes in the two mangrove wetlands (dominance ≥5%)
青梅港 种名 Q1 Q2 Q3 Q4 Q5 Q6 总 Basiria sp. 22.42% 6.90% Daptonema sp.1 5.02% 7.54% 13.21% 5.05% Daptonema sp.2 6.01% 10.69% Daptonema sp.5 6.54% Desmodora sp.1 28.07% 7.63% Haliplectus sp.1 9.09% Linhomoeus sp. 22.55% Neochromadora sp.1 43.89% 28.81% 23.94% 15.95% 22.03% Paracanthonchus sp. 26.36% Paracyatholaimus sp.1 6.99% Paramonohystera sp. 22.27% 13.57% 8.87% Sabatieria sp.1 7.55% T. longicaudata 45.84% 22.55% 42.77% 14.18% Terschellingia sp.2 25.90% 9.43% 5.78% Viscosia sp.1 6.55% 三亚河 种名 S1 S2 S3 S4 S5 S6 总 Chromadorella sp. 5.49% Daptonema sp.1 5.25% 5.49% Eleutherolaimus sp.1 6.40% 25.20% 8.87% 7.05% Haliplectus sp.2 5.00% Metadesmolaimus sp. 8.40% 5.00% Oxystomina sp. 6.25% 7.69% Parodontophora sp. 6.08% 8.94% 7.15% 6.33% Ptycholaimellus sp.2 7.34% 5.21% Sabatieria sp.1 10.23% 25.20% 11.57% 69.11% 11.37% Southerniella sp. 15.00% T. longicaudata 38.36% 25.20% 33.46% 69.11% 46.25% 12.09% 40.09% Terschellingia sp.2 7.05% Theristus sp.1 7.50% Trissonchulus sp.1 6.59% Trissonchulus sp.2 19.78%
下载: 导出CSV
表 5 海洋线虫丰度与环境因子的相关性分析结果
Tab. 5 Results of correlation analysis between abundance of marine nematodes and environmental factors
站点 σi Md OM Chl-a Pheo-a TY Tannin 青梅港 −0.511 0.449 −0.366 0.371 −0.347 −0.52 0.466 三亚河 −0.727 0.729 −0.912* −0.352 −0.851* −0.795 0.452 两个红树林湿地的代表站点 0.294 −0.565 0.434 0.097 −0.233 0.46 0.408 注:*表示在0.05水平(双侧)上显著相关。σi:分选系数;Md:中值粒径;OM:有机质含量;Chl-a:叶绿素含量;Pheo-a:脱镁叶绿酸含量;TY:粉砂黏土含量;Tannin:单宁含量。
下载: 导出CSV
表 6 海洋线虫群落结构DistLM检验结果
Tab. 6 Results of the DIistLM sequential test for marine nematode community structure
环境变量 校准R2 均方 F p Prop Cumul 青梅港 OM − 0.015004 2318.4 0.92609 0.443 0.188 0.188 Md 0.013858 2717 1.1171 0.343 0.22032 0.40831 TY 0.19255 3313.7 1.6639 0.277 0.26871 0.67702 三亚河 Pheo-a 0.30297 4152.6 3.1733 0.011* 0.44237 0.44237 OM 0.45797 2181.6 2.1438 0.116 0.23241 0.67478 TY 0.6141 1603.9 2.2137 0.180 0.17086 0.84564 σi 0.65706 805.18 1.2506 0.445 0.085774 0.93141 青梅港和三亚河代表站点 TY 0.24332 4773.3 2.6078 0.053 0.39466 0.39466 注:Prop:解释差异的比例;F:统计量;Cumul:累计比例。统计上显著的相关性用粗体表示。*表示p < 0.05。
下载: 导出CSV
-
[1] 卢昌义, 林鹏, 叶勇, 等. 全球气候变化对红树林生态系统的影响与研究对策[J]. 地球科学进展, 1995, 10(4): 341−347.Lu Changyi, Lin Peng, Ye Yong, et al. Review on impact of global climate change on mangrove ecosystems and research countermeasure[J]. Advance in Earth Sciences, 1995, 10(4): 341−347. [2] 林鹏. 中国红树林研究进展[J]. 厦门大学学报(自然科学版), 2001, 40(2): 592−603.Lin Peng. A review on the mangrove research in China[J]. Journal of Xiamen University (Natural Science), 2001, 40(2): 592−603. [3] Pinto T K, Austen M C V, Warwick R M, et al. Nematode diversity in different microhabitats in a mangrove region[J]. Marine Ecology, 2013, 34(3): 257−268. doi: 10.1111/maec.12011 [4] 陈映霞. 红树林的环境生态效应[J]. 海洋环境科学, 1995, 14(4): 51−56.Chen Yinxia. Ecologycal effects of the mangrove on the environment[J]. Marine Environmental Science, 1995, 14(4): 51−56. [5] 张志南, 周红. 国际小型底栖生物研究的某些进展[J]. 中国海洋大学学报, 2004, 34(5): 799−806.Zhang Zhinan, Zhou Hong. Some progress on the study of meiofauna[J]. Periodical of Ocean University of China, 2004, 34(5): 799−806. [6] Ridall A, Ingels J. Suitability of free-living marine nematodes as bioindicators: status and future considerations[J]. Frontiers in Marine Science, 2021, 8: 685327. doi: 10.3389/fmars.2021.685327 [7] Moreno M, Semprucci F, Vezzulli L, et al. The use of nematodes in assessing ecological quality status in the Mediterranean coastal ecosystems[J]. Ecological Indicators, 2011, 11(2): 328−336. doi: 10.1016/j.ecolind.2010.05.011 [8] 林鹏, 傅勤. 中国红树林环境生态及经济利用[M]. 北京: 高等教育出版社, 1995.Lin Peng, Fu Qin. Environmental Ecology and Economic Utilization of Mangroves in China[M]. Beijing: Higher Education Press, 1995. [9] 李永翔. 厦门湾不同生境小型底栖动物群落和自由生活海洋线虫分类学研究[D]. 厦门: 集美大学, 2016.Li Yongxiang. Research of meiofauna community and free-living marine nematodes taxonomy from different habitats in Xiamen Bay[D]. Xiamen: Jimei University, 2016. [10] 陈昕韡, 李想, 曾佳丽, 等. 厦门同安湾下潭尾人工红树林湿地小型底栖动物群落结构[J]. 厦门大学学报(自然科学版), 2017, 56(3): 351−358.Chen Xinwei, Li Xiang, Zeng Jiali, et al. Meiofauna communities in artificial mangrove wetland in Xiatanwei of Tong’an Bay, Xiamen[J]. Journal of Xiamen University (Natural Science), 2017, 56(3): 351−358. [11] 曾佳丽, 卓异, 林家逸, 等. 泉州湾洛阳河口红树林湿地自由生活海洋线虫食性研究[J]. 厦门大学学报(自然科学版), 2016, 55(4): 515−521.Zeng Jiali, Zhuo Yi, Lin Jiayi, et al. Feeding habits of free-living marine nematodes in mangrove wetlands in Luoyang Estuary, Quanzhou Bay[J]. Journal of Xiamen University (Natural Science), 2016, 55(4): 515−521. [12] Hu Mingcheng, Guo Yuqing, Shih Y, et al. Comparison of the meiofauna and marine nematode communities before and after removal of Spartina alterniflora in the mangrove wetland of Quanzhou Bay, Fujian Province[J]. Water, 2023, 15(21): 3829. [13] 杨培培, 常瑜, 郭玉清, 等. 福建省红树林湿地海洋线虫的优势属及摄食类型[J]. 海洋学报, 2019, 41(12): 90−102. doi: 10.3969/j.issn.0253-4193.2019.12.009Yang Peipei, Chang Yu, Guo Yuqing, et al. Dominant genera of marine nematodes and their feeding types in several mangrove wetlands of Fujian Province[J]. Haiyang Xuebao, 2019, 41(12): 90−102. doi: 10.3969/j.issn.0253-4193.2019.12.009 [14] 刘梦迪. 福建省红树林湿地常见海洋线虫的分类研究[D]. 厦门: 集美大学, 2018.Liu Mengdi. Research of taxonomy of common free-living marine nematodes in mangrove wetland of Fujian Province[D]. Xiamen: Jimei University, 2018. [15] 傅素晶. 中国东南沿海潮间带小型底栖动物群落生态研究和线虫新种新记录种记述[D]. 厦门: 厦门大学, 2021.Fu Sujing. Ecology of meiofaunal assemblages in the intertidal zone along the southeastern coasts of China and notes of new species and new record species of marine nematodes[D]. Xiamen: Xiamen University, 2021. [16] 傅素晶, 饶义勇, 林和山, 等. 九龙江口潮下带春季小型底栖动物分布及优势类群海洋线虫的群落结构分析[J]. 厦门大学学报(自然科学版), 2023, 62(1): 127−136.Fu Sujing, Rao Yiyong, Lin Heshan, et al. Meiofaunal distribution and assemblages of the dominant group marine nematode in subtidal zone of the Jiulong River estuary in spring[J]. Journal of Xiamen University (Natural Science), 2023, 62(1): 127−136. [17] 肖月平, 郭玉清, 施宜佳. 金门岛红树林湿地海洋线虫群落结构的初步探究[J]. 应用海洋学学报, 2023, 42(3): 402−408.Xiao Yueping, Guo Yuqing, Shi Yijia. Community structures of marine nematodes in Jinmen mangrove wetland[J]. Journal of Applied Oceanography, 2023, 42(3): 402−408. [18] 王玥, 庞小鹏, 郭玉清. 广西北海金海湾红树林湿地海洋线虫群落研究初探[J]. 海洋与湖沼, 2020, 51(3): 583−590. doi: 10.11693/hyhz20191100239Wang Yue, Pang Xiaopeng, Guo Yuqing. Study on marine nematodes community in Jinhai Bay Mangrove wetland, Beihai, Guangxi[J]. Oceanologia et Limnologia Sinica, 2020, 51(3): 583−590. doi: 10.11693/hyhz20191100239 [19] 邹明明, 朱慧兰, 郭玉清. 广西防城港东湾红树林湿地春季小型底栖动物丰度与生物量[J]. 生态学杂志, 2020, 39(6): 1823−1829.Zou Mingming, Zhu Huilan, Guo Yuqing. Abundance and biomass of meiofauna in spring in Dongwan mangrove wetland of Fangchenggang, Guangxi[J]. Chinese Journal of Ecology, 2020, 39(6): 1823−1829. [20] Song Yuanliu, Yan Cunjun, Gao Chunzi, et al. Seasonal distribution of meiofaunal assemblages in the mangrove tidal flat of Futian, Shenzhen, China[J]. Journal of Ocean University of China, 2022, 21(4): 955−964. doi: 10.1007/s11802-022-4869-6 [21] 华尔, 崔春燕, 徐华林, 等. 深圳福田红树林保护区自由生活海洋线虫群落特征研究[J]. 中国海洋大学学报, 2020, 50(9): 46−63.Hua Er, Cui Chunyan, Xu Hualin, et al. Study on the community characteristics of marine nematodes in Futian mangrove reserve, Shenzhen[J]. Periodical of Ocean University of China, 2020, 50(9): 46−63. [22] 潘超, 吴成业, 郭玉清, 等. 湛江特呈岛红树林湿地冬季小型底栖动物和海洋线虫群落的初步研究[J]. 渔业研究, 2020, 42(2): 97−104.Pan Chao, Wu Chengye, Guo Yuqing, et al. Preliminary study on meiofauna and marine nematode community in the mangrove wetland of Techeng Island, Zhanjiang in winter[J]. Journal of Fisheries Research, 2020, 42(2): 97−104. [23] 潘超, 郭玉清, 陈芳. 高桥红树林湿地冬季海洋线虫群落结构[J]. 集美大学学报(自然科学版), 2022, 27(1): 17−23.Pan Chao, Guo Yuqing, Chen Fang. Community structure of marine nematodes in winter in Gaoqiao mangrove wetland[J]. Journal of Jimei University (Natural Science), 2022, 27(1): 17−23. [24] 肖月平. 台湾省红树林湿地海洋线虫的群落结构与分类研究[D]. 厦门: 集美大学, 2023.Xiao Yueping. Study on community structure and classification of marine nematodes in mangrove wetlands in Taiwan Province[D]. Xiamen: Jimei University, 2023. [25] 邹明明, 郭玉清, 甘加成. 秋季海南省新英港红树林湿地的海洋线虫群落结构研究[J]. 中国海洋大学学报, 2022, 52(5): 44−54.Zou Mingming, Guo Yuqing, Gan Jiacheng. Studies on the community structure of Xinying Port (Hainan Province) mangrove wetland marine nematodes in autumn[J]. Periodical of Ocean University of China, 2022, 52(5): 44−54. [26] 宿少锋, 林之盼, 王小燕, 等. 海南红树林湿地生态系统退化程度评价指标体系构建研究[J]. 林业资源管理, 2022(6): 54−60.Su Shaofeng, Lin Zhipan, Wang Xiaoyan, et al. Research on the construction of the evaluation index system for the degradation degree of mangrove wetland ecosystem in Hainan[J]. Forest Resources Management, 2022(6): 54−60. [27] 黄良民, 张偲, 王汉奎, 等. 三亚湾生态环境与生物资源[M]. 北京: 科学出版社, 2007.Huang Liangmin, Zhang Si, Wang Hankui, et al. Ecological Environment and Bio-Resources for Sanya Bay, Hainan Island, China[M]. Beijing: Science Press, 2007. [28] 姚轶锋, 廖文波, 宋晓彦, 等. 海南三亚铁炉港红树林资源现状与保护[J]. 海洋通报, 2010, 29(2): 150−155. doi: 10.3969/j.issn.1001-6392.2010.02.005Yao Yifeng, Liao Wenbo, Song Xiaoyan, et al. Current status and conservation of mangrove resources in Tielu Harbor, Sanya, Hainan[J]. Marine Science Bulletin, 2010, 29(2): 150−155. doi: 10.3969/j.issn.1001-6392.2010.02.005 [29] 符国瑷, 黎军. 海南三亚市红树林植被调查初报[J]. 海南大学学报(自然科学版), 2000, 18(3): 287−292.Fu Guoai, Li Jun. Preliminary report of investigation regarding mangrove of Shanya City, Hainan[J]. Journal of Hainan University Natural Science, 2000, 18(3): 287−292. [30] 刘均玲, 黄勃, 梁志伟. 东寨港红树林小型底栖动物的密度和生物量研究[J]. 海洋学报, 2013, 35(2): 187−192. doi: 10.3969/j.issn.0253-4193.2013.02.020Liu Junling, Huang Bo, Liang Zhiwei. Study on abundance and biomass of benthic meiofauna in mangrove of Dongzhai Bay[J]. Haiyang Xuebao, 2013, 35(2): 187−192. doi: 10.3969/j.issn.0253-4193.2013.02.020 [31] 刘均玲, 袁超, 何永姑, 等. 东寨港红树林小型底栖动物丰度与Chla、有机质的相关性[J]. 生态学报, 2019, 39(1): 185−191.Liu Junling, Yuan Chao, He Yonggu, et al. Correlation of meiofauna abundance with chlorophyll a and organic matter of Dongzhai Harbor Mangrove[J]. Acta Ecologica Sinica, 2019, 39(1): 185−191. [32] 袁超. 海南东寨港红树林小型底栖动物分布与主要环境因子的相关性研究[D]. 海口: 海南大学, 2017.Yuan Chao. Distribution of meiofauna and its correlation with main environmental factors in Dongzhai port Mangrove of Hainan province[D]. Haikou: Hainan University, 2017. [33] 何永姑, 刘均玲, 袁超, 等. 不同季节东寨港红树林沉积物小型底栖动物的分布特征[J]. 海洋科学进展, 2019, 37(4): 681−688.He Yonggu, Liu Junling, Yuan Chao, et al. Characteristics of meiofauna distribution in sediments of Dongzhai Bay Mangrove in different seasons[J]. Advances in Marine Science, 2019, 37(4): 681−688. [34] 王娟, 宋迎春, 张烨, 等. 东寨港红树林凋落叶附生小型底栖动物的群落特征及影响因素[J]. 海洋科学, 2023, 47(9): 28−39. doi: 10.11759/hykx20230225001Wang Juan, Song Yingchun, Zhang Ye, et al. Community characteristics and factors affecting meiofauna on mangrove leaf litter from Dongzhai Harbor[J]. Marine Sciences, 2023, 47(9): 28−39. doi: 10.11759/hykx20230225001 [35] 钟才荣, 林贵生, 陈元海, 等. 三亚清梅港红树林群落特征调查[J]. 热带林业, 2009, 37(3): 45-47.Zhong Cairong, Lin Guisheng, Chen Yuanhai, et al. Investigation on mangrove community characteristics in Qingmei Harbor, Sanya[J]. Tropical Forestry, 2009, 37(3): 45−47. [36] 王丽荣, 李贞, 蒲杨婕, 等. 近50年海南岛红树林群落的变化及其与环境关系分析——以东寨港、三亚河和青梅港红树林自然保护区为例[J]. 热带地理, 2010, 30(2): 114−120. doi: 10.3969/j.issn.1001-5221.2010.02.003Wang Lirong, Li Zhen, Pu Yangjie, et al. Analysis on the relationship between mangrove and environment change in Hainan Island in the past 50 years: a case study of Dongzhai Harbor, Sanya River and Qingmei Harbor Mangrove nature protection area[J]. Tropical Geography, 2010, 30(2): 114−120. doi: 10.3969/j.issn.1001-5221.2010.02.003 [37] 王冰鑫. 三亚河红树林生态系统群落结构调查及碳储量研究[D]. 三亚: 海南热带海洋学院, 2022.Wang Bingxin. Investigation on community structure and carbon storage of mangrove ecosystem in Sanya River[D]. Sanya: Hainan Tropical Ocean University, 2022. [38] Wieser W. Die Beziehung zwischen Mundhöhlengestalt, Ernährungsweise und Vorkommen bei freilebenden marinen Nematoden[J]. Arkiv fö r Zoologi, 1953, 4(26): 439−484. [39] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. 海洋监测规范: 第5部分 沉积物分析: GB 17378.5−2007[S]. 北京: 中国标准出版社, 2008.General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, Standardization Administration of the People’s Republic of China. The specification for marine monitoring—Part 5: sediment analysis GB 17378.5−2007[S]. Beijing: Standards Press of China, 2008. [40] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. 海洋调查规范: 第6部分 海洋生物调查: GB/T 12763.6—2007[S]. 北京: 中国标准出版社, 2008.General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China, Standardization Administration of the People's Republic of China. The specifications for oceanographic survey—Part 6: marine biological survey GB/T 12763.6−2007[S]. Beijing: Standards Press of China, 2008. [41] Allen S E, Grimshaw H M, Parkinson J A, et al. Chemical Analysis of Ecological Materials[M]. Oxford: Blackwell Scientific Publications, 1974. [42] 周红, Uddin S M D, 黄梦娇, 等. 南海北部陆架和陆坡区小型底栖动物群落的比较研究[J]. 海洋与湖沼, 2020, 51(3): 555−563. doi: 10.11693/hyhz20191100228Zhou Hong, Uddin S M D, Huang Mengjiao, et al. Comparison in meiofaunal communities between shelf and slope of the northern South China Sea[J]. Oceanologia et Limnologia Sinica, 2020, 51(3): 555−563. doi: 10.11693/hyhz20191100228 [43] Soetaert K, Franco M, Lampadariou N, et al. Factors affecting nematode biomass, length and width from the shelf to the deep sea[J]. Marine Ecology Progress Series, 2009, 392: 123−132. doi: 10.3354/meps08202 [44] 安嘉然, 臧金娇, 刘金福. 三亚地区红树林湿地及其栖息地的发展变化现状与对策[J]. 武夷科学, 2013, 29(1): 30−37. doi: 10.3969/j.issn.1001-4276.2013.01.005An Jiaran, Zang Jinjiao, Liu Jinfu. The development status and countermeasures of mangrove wetland in Sanya area[J]. Wuyi Science Journal, 2013, 29(1): 30−37. doi: 10.3969/j.issn.1001-4276.2013.01.005 [45] Hua Er, Zhu Yanmei, Huang Deming, et al. Are free-living nematodes effective environmental quality indicators? Insights from Bohai Bay, China[J]. Ecological Indicators, 2021, 127: 107756. doi: 10.1016/j.ecolind.2021.107756 [46] Moens T, Braeckman U, Derycke S, et al. Ecology of free-living marine nematodes[J]. Handbook of Zoology: Gastrotricha, Cycloneuralia and Gnathifera, 2013, 2: 109−152. [47] 吴昌文, 李志国, 夏武强. 小型底栖动物(Meiofauna)研究概况[J]. 现代渔业信息, 2008, 23(3): 9−12.Wu Changwen, Li Zhiguo, Xia Wuqiang. Research outling on Meiofauna[J]. Modern Fisheries Information, 2008, 23(3): 9−12. [48] 刘清河, 马林, 李新正. 东海北部小型底栖动物群落对径流及黑潮暖流入侵的响应[J]. 海洋学报, 2020, 42(2): 52−64. doi: 10.3969/j.issn.0253-4193.2020.02.006Liu Qinghe, Ma Lin, Li Xinzheng. The communities of meiofauna in the northern East China Sea and their responses to runoff and the intrusion of Kuroshio Current[J]. Haiyang Xuebao, 2020, 42(2): 52−64. doi: 10.3969/j.issn.0253-4193.2020.02.006 [49] Middelburg J J, Nieuwenhuize J, Slim F J, et al. Sediment biogeochemistry in an East African mangrove forest (Gazi Bay, Kenya)[J]. Biogeochemistry, 1996, 34(3): 133−155. [50] Marchand C, Baltzer F, Lallier-Vergès E, et al. Pore-water chemistry in mangrove sediments: relationship with species composition and developmental stages (French Guiana)[J]. Marine Geology, 2004, 208(2/4): 361−381. [51] 王爱军, 高抒, 贾建军. 互花米草对江苏潮滩沉积和地貌演化的影响[J]. 海洋学报, 2006, 28(1): 92−99. doi: 10.3321/j.issn:0253-4193.2006.01.013Wang Aijun, Gao Shu, Jia Jianjun. Impact of Spartina alterniflora on sedimentary and morphological evolution of tidal salt marshes of Jiangsu, China[J]. Haiyang Xuebao, 2006, 28(1): 92−99. doi: 10.3321/j.issn:0253-4193.2006.01.013 [52] 袁兴中, 陆健健. 潮滩微地貌元素——“生物结构”与小型底栖动物的空间分布[J]. 生态学杂志, 2003, 22(6): 124−126. doi: 10.3321/j.issn:1000-4890.2003.06.025Yuan Xingzhong, Lu Jianjian. Micro-topographical element—“Biogenic Structure” and spatial distribution of meiofauna on the tidal flat[J]. Chinese Journal of Ecology, 2003, 22(6): 124−126. doi: 10.3321/j.issn:1000-4890.2003.06.025 [53] Chinnadurai G, Fernando O J. Meiofauna of mangroves of the southeast coast of India with special reference to the free-living marine nematode assemblage[J]. Estuarine, Coastal and Shelf Science, 2007, 72(1/2): 329−336. [54] Coull B C. Role of meiofauna in estuarine soft-bottom habitats[J]. Australian Journal of Ecology, 1999, 24(4): 327−343. doi: 10.1046/j.1442-9993.1999.00979.x [55] 郭玉清. 厦门凤林红树林湿地自由生活海洋线虫群落的研究[J]. 海洋学报, 2008, 30(4): 147−153. doi: 10.3321/j.issn:0253-4193.2008.04.018Guo Yuqing. The study on the community of free-living marine nematodes in Fenglin mangrove wetlands, Xiamen, China[J]. Haiyang Xuebao, 2008, 30(4): 147−153. doi: 10.3321/j.issn:0253-4193.2008.04.018 [56] Ghosh M, Mandal S. Does vertical distribution of meiobenthic community structure differ among various mangrove habitats of Sundarban Estuarine System?[J]. Regional Studies in Marine Science, 2019, 31: 100778. doi: 10.1016/j.rsma.2019.100778 [57] 何蕾, 华尔, 刘晓收, 等. 夏、秋季渤海小型底栖动物类群组成及分布特征[J]. 生态学报, 2019, 39(8): 2794−2805.He Lei, Hua Er, Liu Xiaoshou, et al. Meiofauna assemblage composition and distribution in the Bohai Sea during summer and autumn[J]. Acta Ecologica Sinica, 2019, 39(8): 2794−2805. [58] Warwick R M. Nematode associations in the Exe estuary[J]. Journal of the Marine Biological Association of the United Kingdom, 1971, 51(2): 439−454. [59] Heip C H R, Vincx M, Vranken G. The ecology of marine nematodes[J]. Oceanography and Marine Biology, 1985, 23: 399−489. [60] 袁俏君, 苗素英, 李恒翔, 等. 珠江口水域夏季小型底栖生物群落结构[J]. 生态学报, 2012, 32(19): 5962−5971.Yuan Qiaojun, Miao Suying, Li Hengxiang, et al. Analysis of the meiobenthic community in the Pearl River Estuary in summer[J]. Acta Ecologica Sinica, 2012, 32(19): 5962−5971. [61] Montagna P A, Kalke R D. The effect of freshwater inflow on meiofaunal and macrofaunal populations in the Guadalupe and Nueces Estuaries, Texas[J]. Estuaries, 1992, 15(3): 307−326. [62] Ingels J, dos Santos G, Hicks N, et al. Short-term CO2 exposure and temperature rise effects on metazoan meiofauna and free-living nematodes in sandy and muddy sediments: results from a flume experiment[J]. Journal of Experimental Marine Biology and Ecology, 2018, 502: 211−226. doi: 10.1016/j.jembe.2017.07.012 [63] 王丽荣, 李贞, 蒲杨婕, 等. 海南东寨港、三亚河和青梅港红树林群落健康评价[J]. 热带海洋学报, 2011, 30(2): 81−86. doi: 10.3969/j.issn.1009-5470.2011.02.012Wang Lirong, Li Zhen, Pu Yangjie, et al. Health evaluation of mangrove community in Dongzhaigang, Sanyahe, and Qingmeigang Mangrove Nature Reserves of Hainan Province[J]. Journal of Tropical Oceanography, 2011, 30(2): 81−86. doi: 10.3969/j.issn.1009-5470.2011.02.012 -

计量
- 文章访问数: 295
- HTML全文浏览量: 121
- PDF下载量: 19
- 被引次数: 0
