

Preliminary study on the extinction of Gymnodinium catenatum bloom and its relationship with resting cysts
-
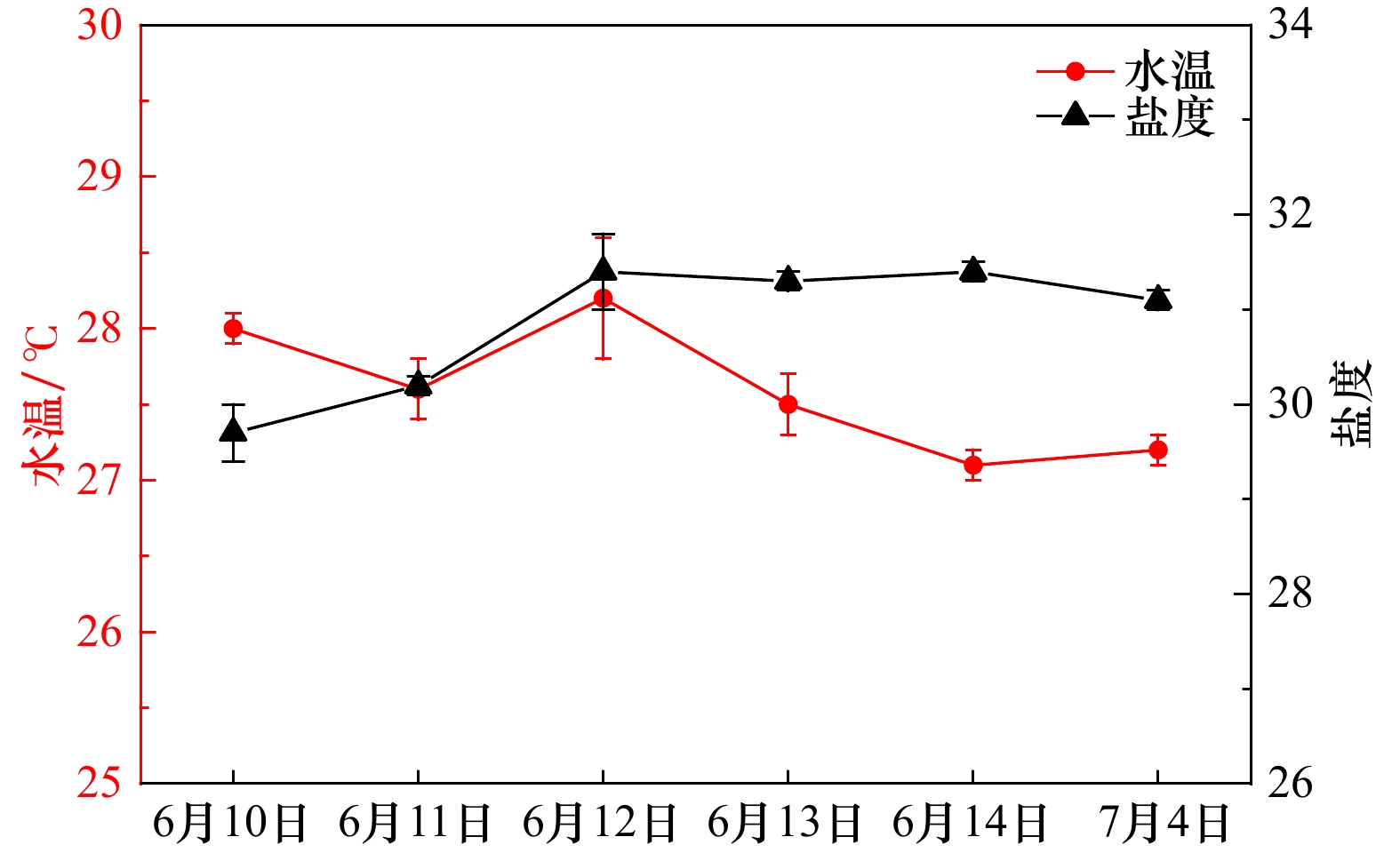
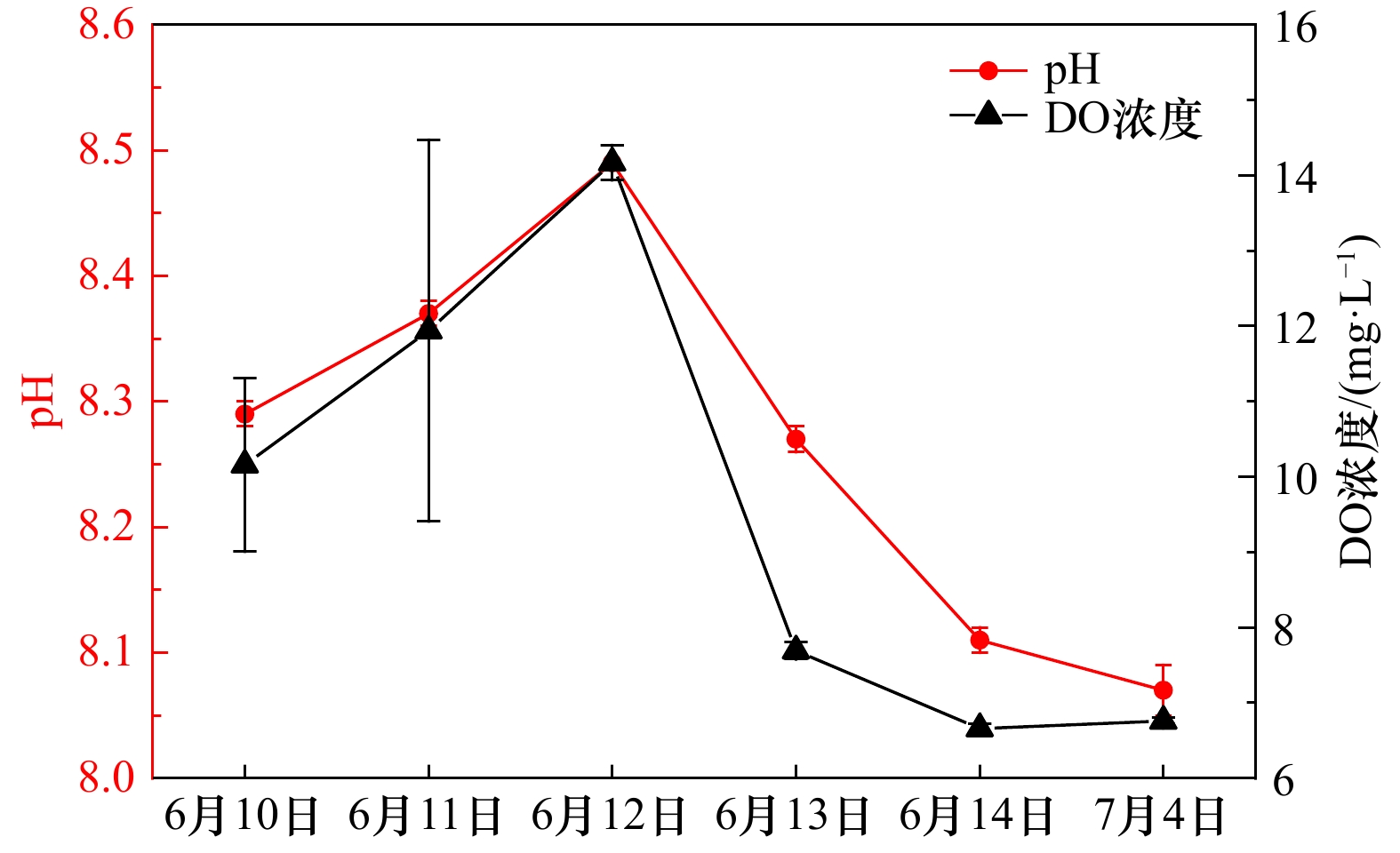
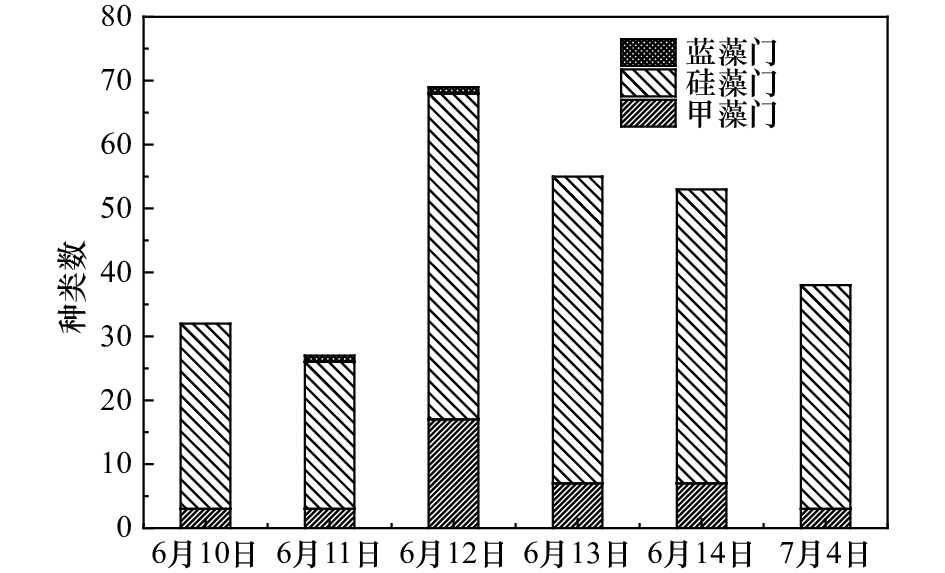
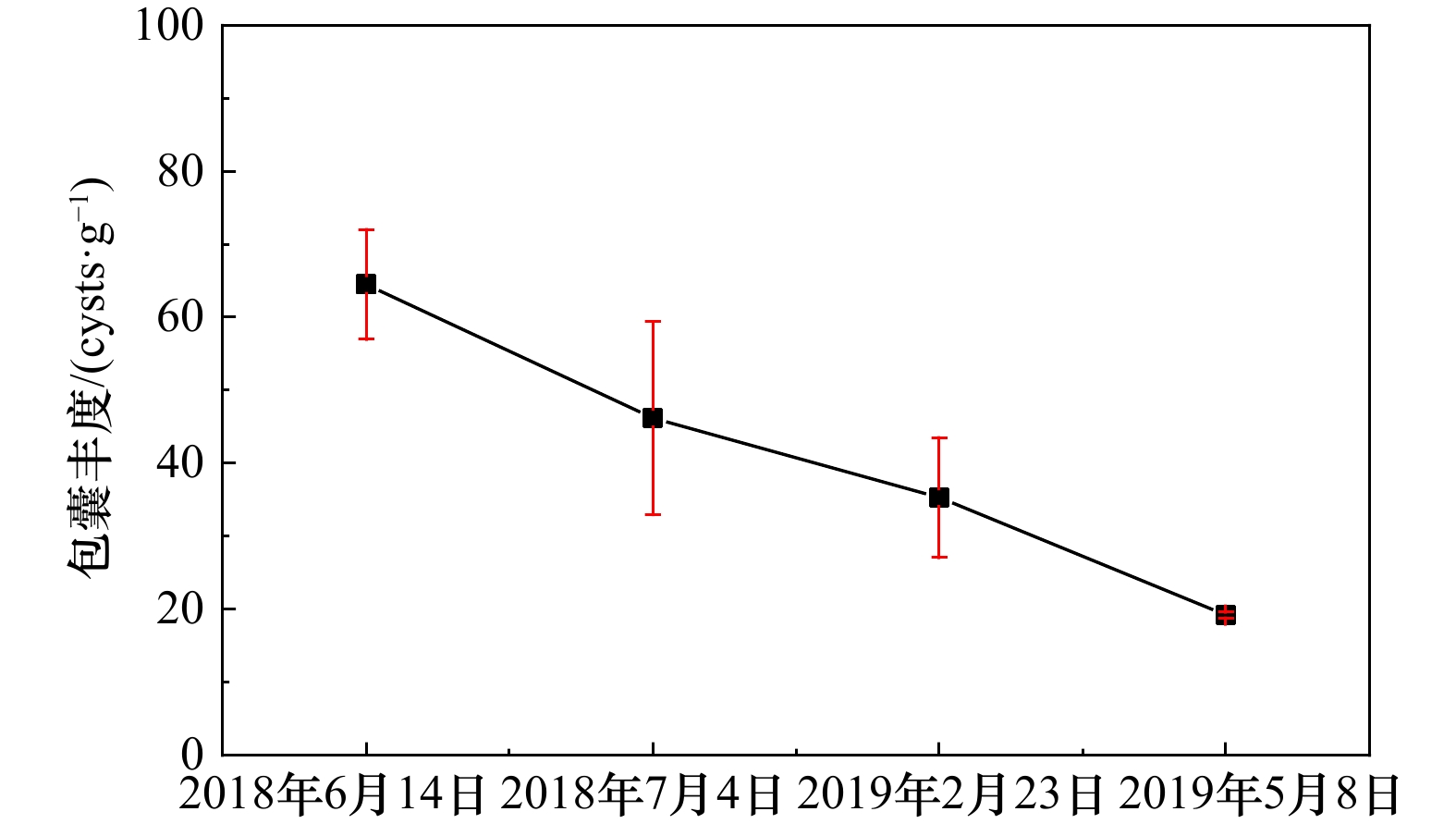
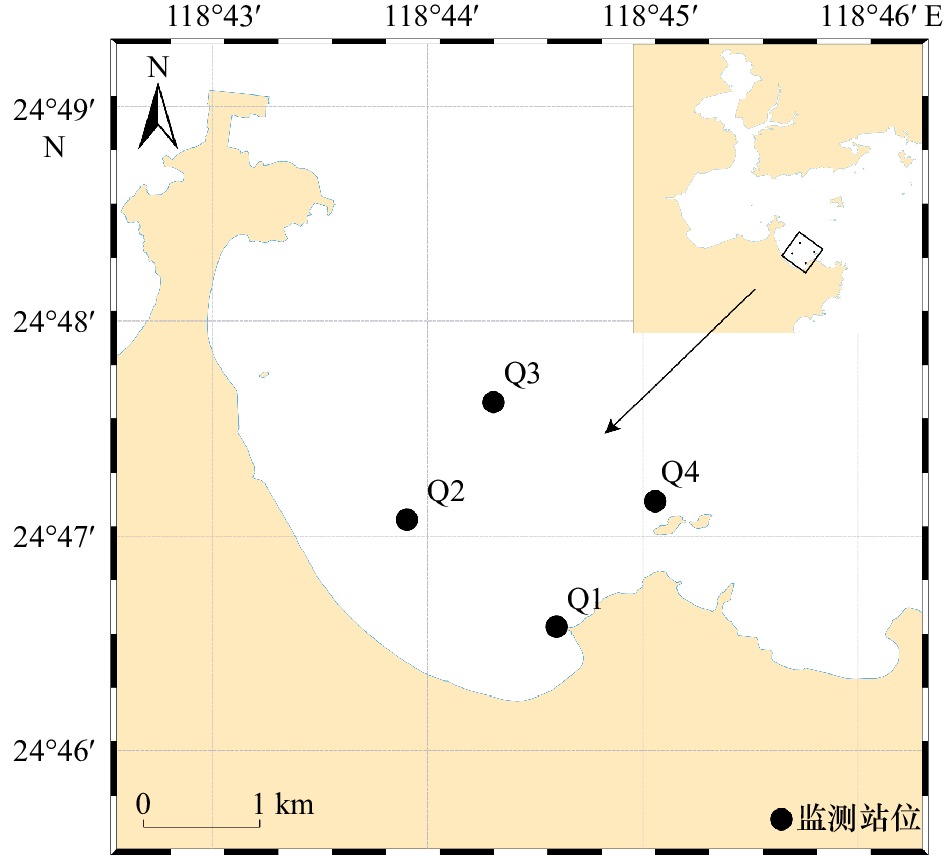
摘要: 本文对2018年6月10−14日在泉州湾海域发生的一次链状裸甲藻(Gymnodinium catenatum)赤潮的消亡过程进行研究。在现场跟踪监测,共设置4个站位,进行6个航次海洋环境及浮游植物的调查。结合本文对泉州湾海域甲藻包囊的调查研究,探索休眠包囊与赤潮生消过程的关系。研究结果显示,赤潮发生时海况良好,水温和盐度分别介于27.1~28.2℃和29.7~31.4之间,活性磷酸盐和无机氮浓度分别介于未检出至0.045 4 mg/L和0.050~0.281 mg/L之间,赤潮生物密度与营养盐浓度没有呈现显著的相关关系,大风大浪是导致赤潮消亡的重要原因;赤潮消亡过程中共鉴定出浮游植物3门48属100种,硅藻门种类最多;赤潮消亡期间浮游植物群落结构发生变化,前期链状裸甲藻占绝对优势,最高密度达(7.02±0.11)×106 cells/L,后期中肋骨条藻(Skeletonema costatum)和旋链角毛藻(Chaetoceros curvisetus)代替链状裸甲藻成为优势种,且浮游植物种类数明显增加;同时在泉州湾海域共鉴定甲藻包囊5大类37种和1种未定种,平均丰度为574 cysts/g,以异养型甲藻包囊为主;休眠包囊在赤潮生消过程中起着重要作用,赤潮消亡时形成休眠包囊,包囊数量会随着时间呈下降趋势,但仍有萌发的可能,具有重新暴发赤潮的潜在性,需要引起重视。Abstract: During June 10 to 14, 2018, the extinction of Gymnodinium catenatum bloom in the Quanzhou Bay was studied in this paper. In the field investigation, a total of 4 stations were set up to monitor the marine environment factors and phytoplankton within 6 times. Combined with the investigation of dinoflagellate cysts in the Quanzhou Bay, the relationship of dinoflagellate cysts and the evolution of bloom was explored. The results showed that when the bloom occurred, the sea state was good and the water temperature and salinity were 27.1℃ to 28.2℃ and 29.7 to 31.4 respectively. The concentration of PO4-P and inorganic nitrogen were between below detectable limit to 0.0454 mg/L and 0.050 mg/L to 0.281 mg/L respectively. There was no significant correlation between the density of bloom biology and nutrients. The strong winds and waves were the important factors for the extinction of bloom. One hundred species belonging to 48 genera and 3 phyla of phytoplankton were identified in the extinction of bloom, and Bacillariophyta was the most phylum. The community structure of phytoplankton changed during the extinction of bloom, Gymnodinium catenatum was the absolute dominant, the maximum density of which reached (7.02±0.11)×106 cells/L. In the declination of the bloom, Skeletonema costatum and Chaetoceros curvisetus dominated, and the number of phytoplankton species increased significantly. At the same time, 37 species of dinoflagellate cysts in 5 groups and 1 undetermined species were identified in the Quanzhou Bay, and the average abundance of which was 574 cysts/g, mainly dominated by heterotrophic dinoflagellate cysts. The resting cysts played an important role in the evolution of bloom. They occurred when the bloom dissipated and decreased with time. However, they still could germinate and had the potential to re-outbreak and form bloom, which should raise concern.
-
Key words:
- Quanzhou Bay /
- Gymnodinium catenatum /
- red tide /
- resting cysts
-
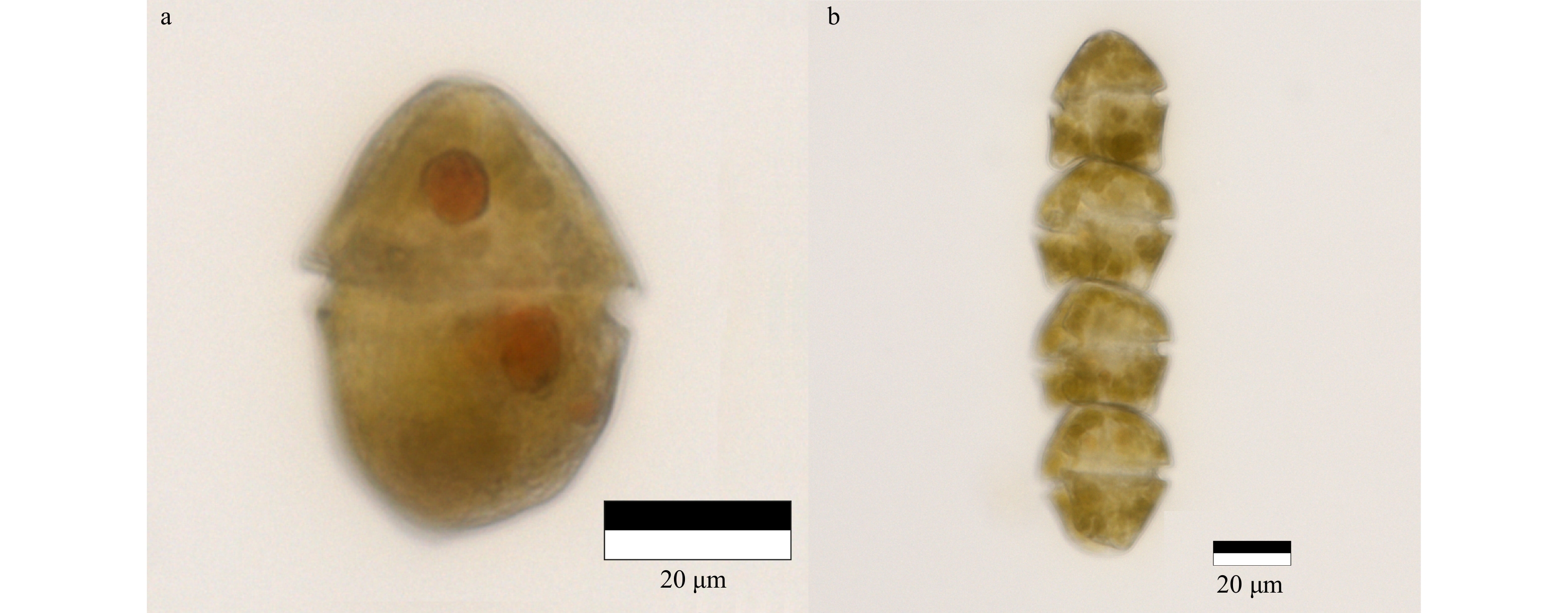
图 2 链状裸甲藻单细胞(a)及链状群体(b)
Fig. 2 Gymnodinium catenatum monoplast (a) and catenoid colony (b)
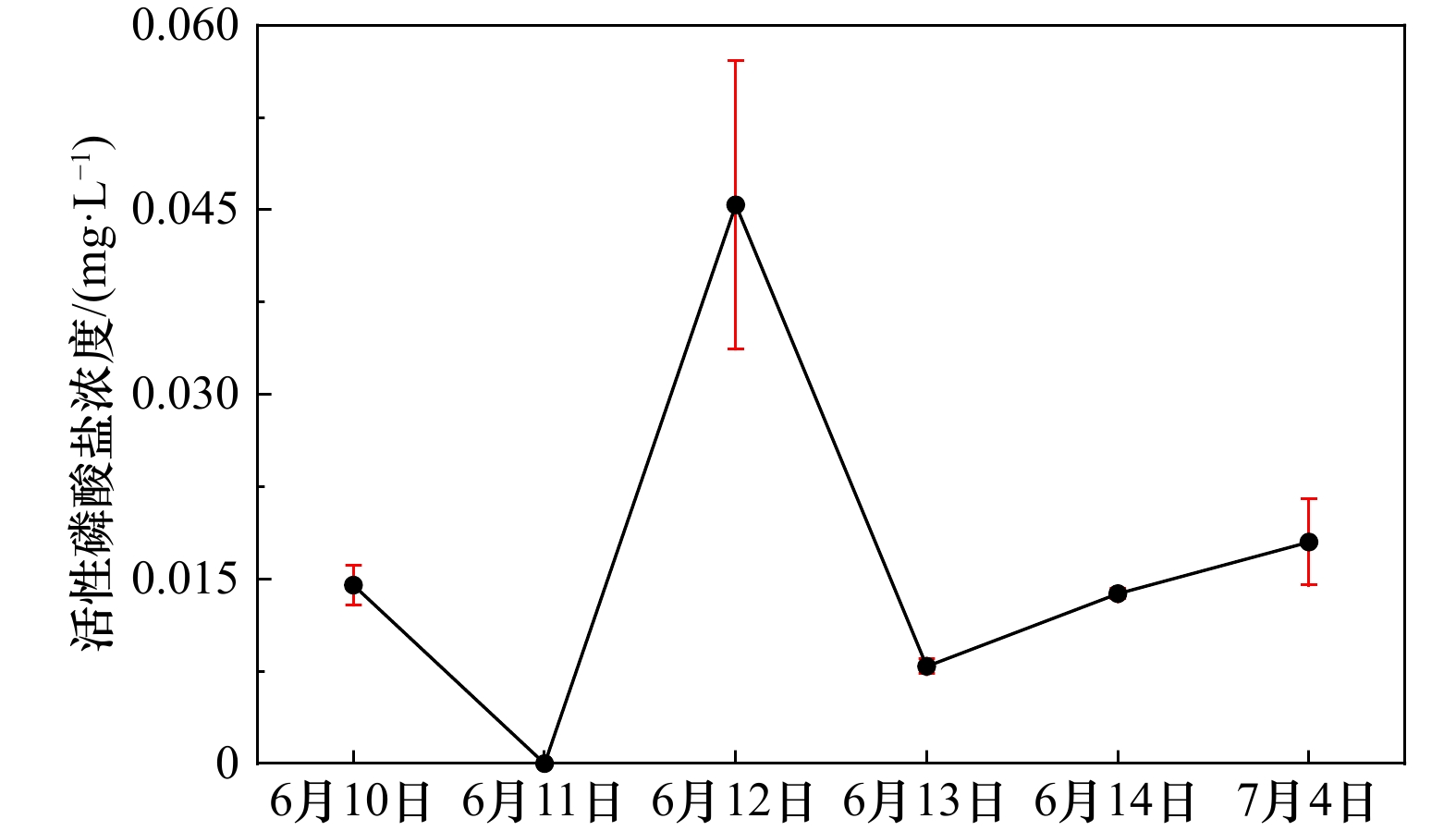
图 5 赤潮期间活性磷酸盐(PO4-P)浓度的变化
Fig. 5 Variations of active phosphate (PO4-P) concentration during red tide
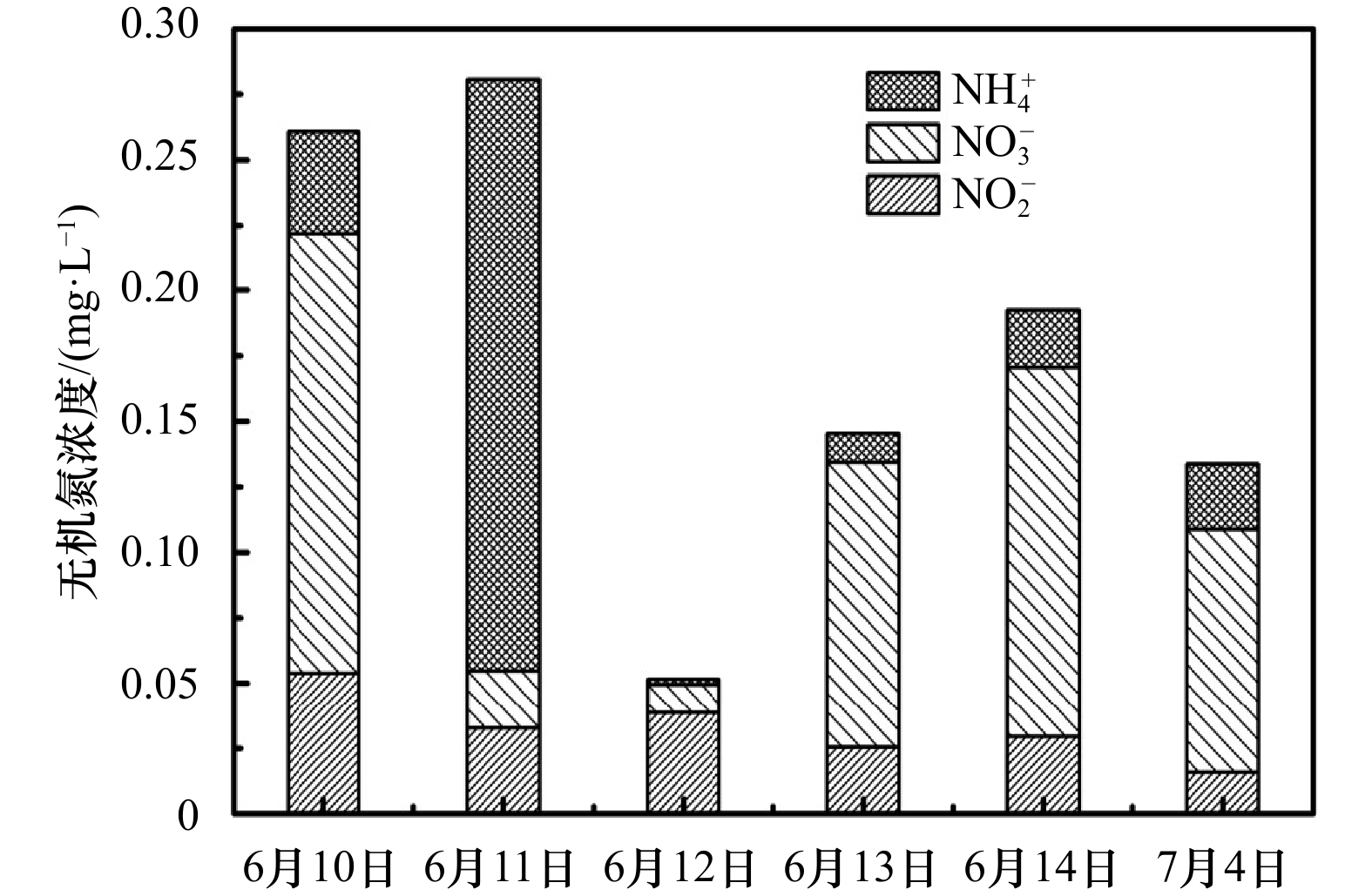
图 6 赤潮期间无机氮浓度的变化
Fig. 6 Variations of inorganic nitrogen concentration during red tide
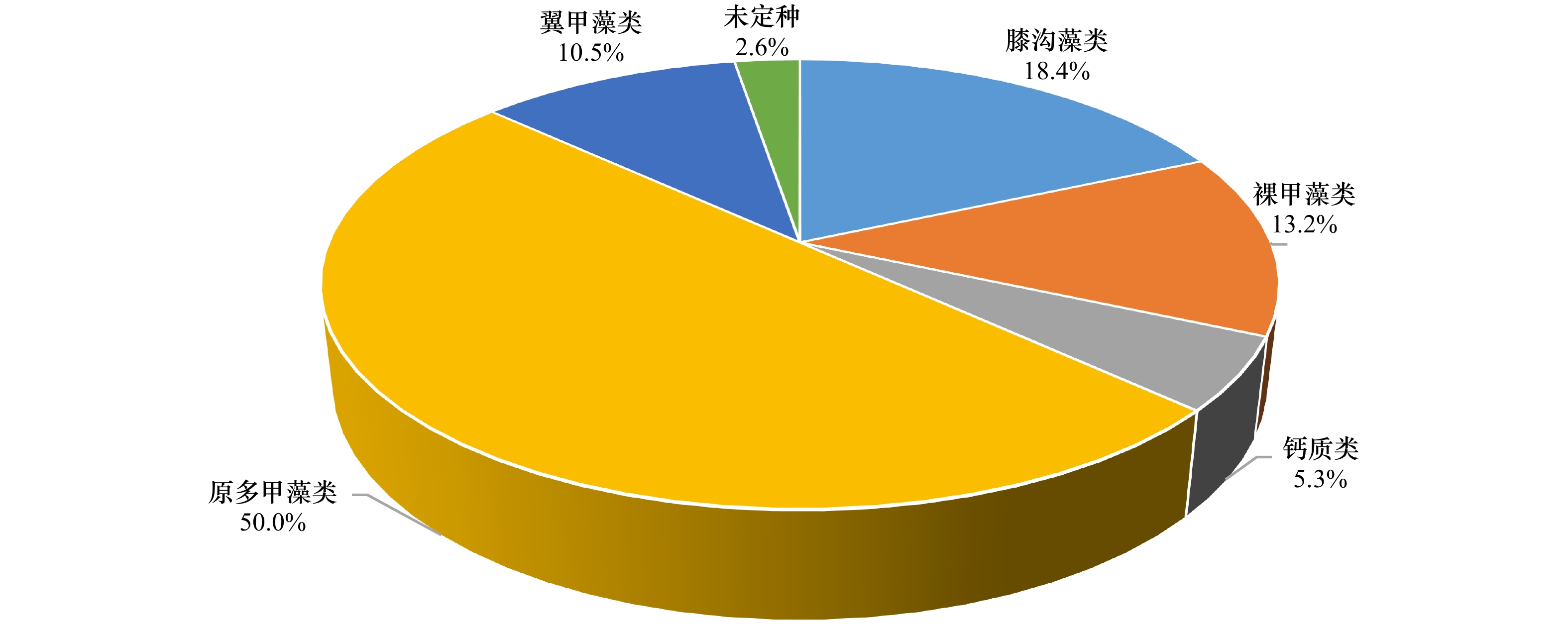
图 7 泉州湾海域甲藻包囊种类组成
Fig. 7 Species composition of dinoflagellate cysts in the Quanzhou Bay
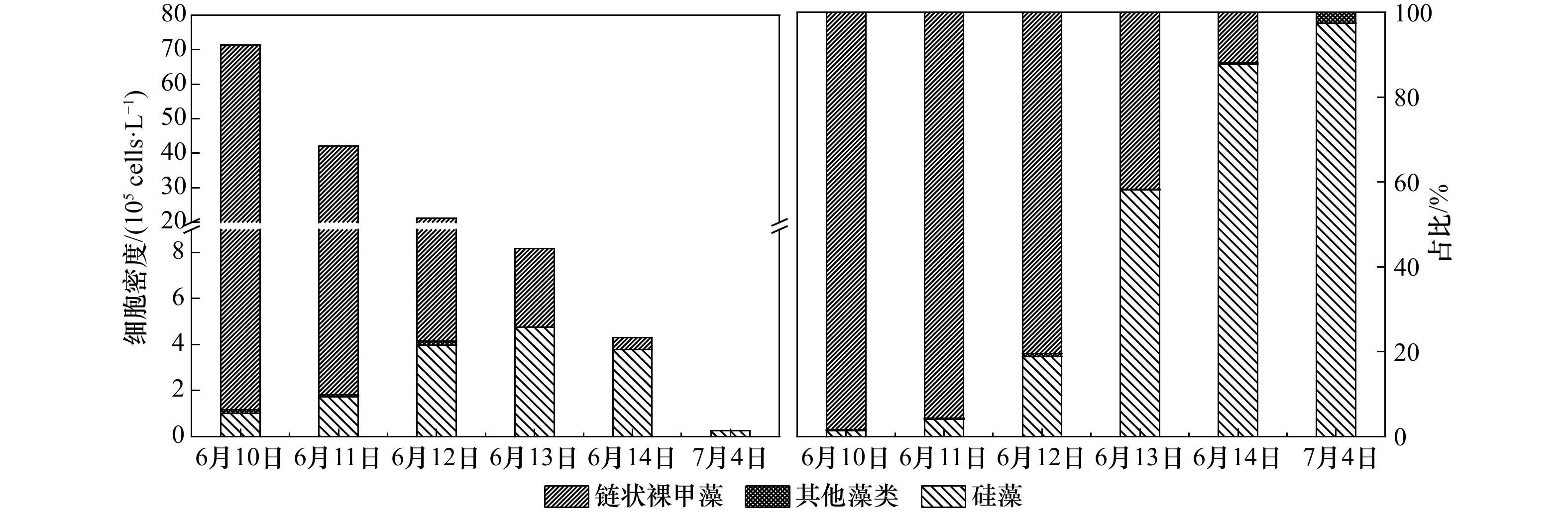
图 9 赤潮期间浮游植物细胞密度及其所占比率的变化
Fig. 9 Variations of phytoplankton cell density and its proportion during red tide
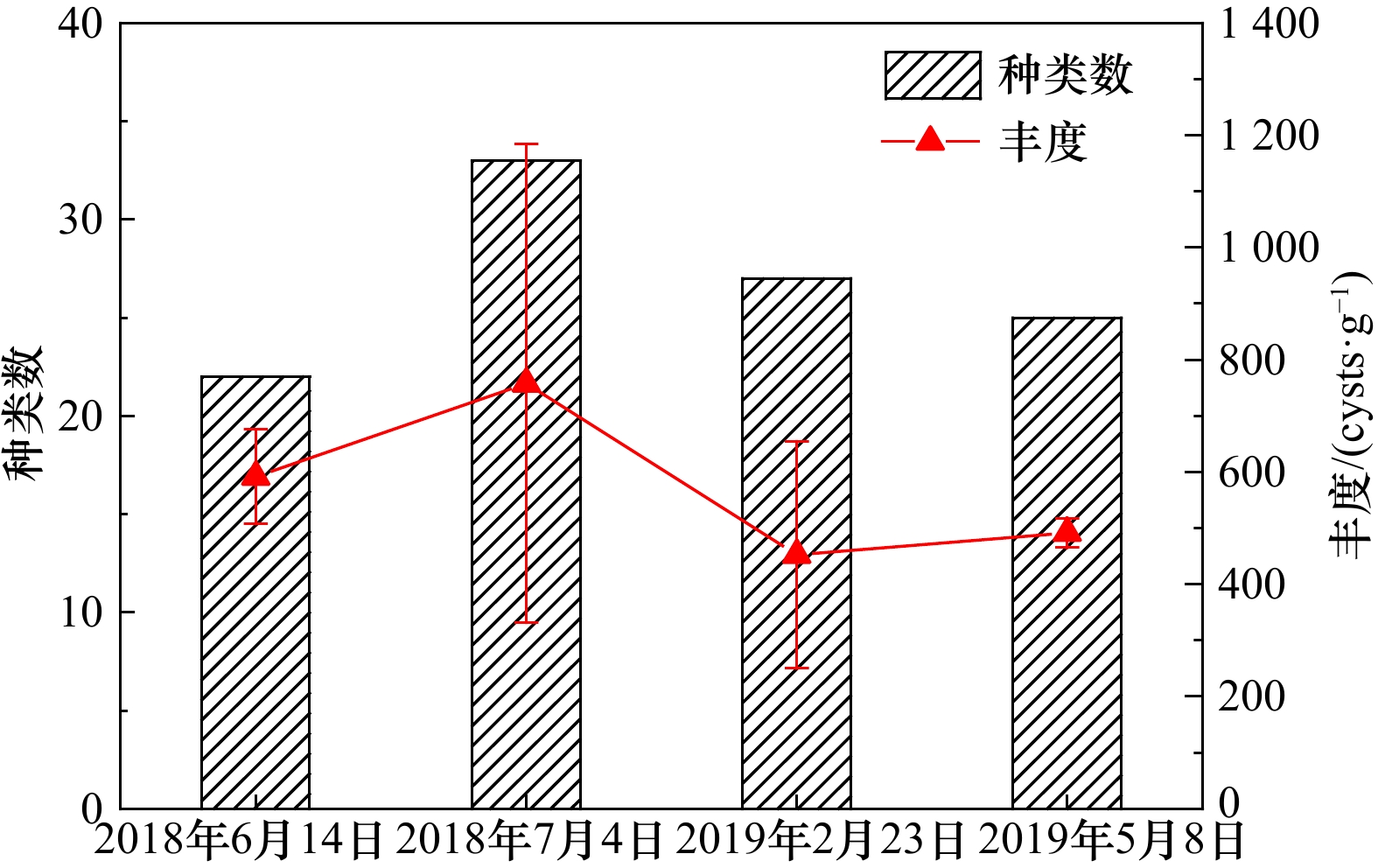
图 10 泉州湾海域甲藻包囊种类数和丰度的变化
Fig. 10 Variations of species number and abundance of dinoflagellate cysts in the Quanzhou Bay

图 11 链状裸甲藻空包囊(a)和活体包囊(b)
Fig. 11 Gymnodinium catenatum empty cysts(a)and live cysts(b)
-
[1] 李冠国, 范振刚. 海洋生态学[M]. 2版. 北京: 高等教育出版社, 2011: 343.Li Guanguo, Fan Zhengang. Marine Ecology[M]. 2nd ed. Beijing: Higher Education Press, 2011: 343. [2] 李雪丁. 近20年福建沿海赤潮的发生特点[J]. 海洋环境科学, 2021, 40(4): 601−610. doi: 10.12111/j.mes.20200120Li Xueding. Occurrence characteristics of the red tide in Fujian coastal waters during the last two decades[J]. Marine Environmental Science, 2021, 40(4): 601−610. doi: 10.12111/j.mes.20200120 [3] 高养春, 董燕红, 李海涛, 等. 有害甲藻包囊的分类鉴定研究进展[J]. 生物安全学报, 2016, 25(4): 238−254. doi: 10.3969/j.issn.2095-1787.2016.04.002Gao Yangchun, Dong Yanhong, Li Haitao, et al. Research progress on identification of harmful dinoflagellate cysts: a review[J]. Journal of Biosafety, 2016, 25(4): 238−254. doi: 10.3969/j.issn.2095-1787.2016.04.002 [4] 丁光茂, 张树峰. 2012年三沙湾米氏凯伦藻赤潮的生态特征及成因分析[J]. 海洋学报, 2018, 40(6): 104−112.Ding Guangmao, Zhang Shufeng. Ecological characteristics and the causes of Karenia mikimotoi bloom in the Sansha Bay in 2012[J]. Haiyang Xuebao, 2018, 40(6): 104−112. [5] 郭皓, 丁德文, 林凤翱, 等. 近20 a我国近海赤潮特点与发生规律[J]. 海洋科学进展, 2015, 33(4): 547−558. doi: 10.3969/j.issn.1671-6647.2015.04.013Guo Hao, Ding Dewen, Lin Fengxiang, et al. Characteristics and patterns of red tide in China coastal waters during the last 20 a[J]. Advances in Marine Science, 2015, 33(4): 547−558. doi: 10.3969/j.issn.1671-6647.2015.04.013 [6] 张文. 不同环境因子对有害赤潮生物链状裸甲藻的生长和产毒的影响[D]. 广州: 暨南大学, 2009.Zhang Wen. Effect of different environment factors on the growth and toxin production of toxic dinoflagellate Gymnodinium catenatum Graham[D]. Guangzhou: Jinan University, 2009. [7] 王朝晖. 中国典型近海海域甲藻包囊分布及其与富营养化和赤潮生消关系研究[D]. 广州: 暨南大学, 2007.Wang Zhaohui. Distribution of dinoflagellate cysts in Chinese typical coastal areas and its relationships with eutrophication and algal bloom[D]. Guangzhou: Jinan University, 2007. [8] 陈火荣. 2017年福建海域链状裸甲藻赤潮事件应急处置与思考[J]. 渔业研究, 2018, 40(4): 308−314.Chen Huorong. Emergency treatment and reflection of red tide event of Gymnodinium catenatum in Fujian sea area in 2017[J]. Journal of Fisheries Research, 2018, 40(4): 308−314. [9] 颜利, 蒋金龙, 詹兴旺, 等. 泉州湾表层水主要化学要素含量和富营养化指数的时空变化特征[J]. 台湾海峡, 2012, 31(4): 459−465.Yan Li, Jiang Jinlong, Zhan Xingwang, et al. Characters of spatiotemporal variations of chemical elements content and eutrophication index in surface water of Quanzhou Bay[J]. Journal of Oceanography in Taiwan Strait, 2012, 31(4): 459−465. [10] 国家质量监督检验检疫总局. GB/T 12763−2007 海洋调查规范[S]. 北京: 中国标准出版社, 2007.General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China. GB/T 12763−2007 Specifications for oceanographic survey[S]. Beijing: China Standards Press, 2007. [11] 国家质量监督检验检疫总局. GB 17378−2007 海洋监测规范[S]. 北京: 中国标准出版社, 2007.General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China. GB 17378−2007 The specification for marine monitoring[S]. Beijing: China Standards Press, 2007. [12] 王朝晖. 中国沿海甲藻包囊与赤潮研究[M]. 北京: 海洋出版社, 2007.Wang Zhaohui. Study of Dinoflagellate Cysts and Red Tide in Coastal Areas of China Sea[M]. Beijing: China Ocean Press, 2007. [13] 陆欣鑫. 华南沿海典型海域甲藻包囊地理分布及环境指示作用研究[D]. 广州: 暨南大学, 2017.Lu Xinxin. Biogeological distribution of dinoflagellate cysts in surface sediments from southern Chinese coast and their roles as a signal of environmental changes[D]. Guangzhou: Jinan University, 2017. [14] 郭皓. 我国海域赤潮甲藻包囊形态与分布特征研究[D]. 大连: 大连海事大学, 2016.Guo Hao. Studies on the characteristics of the morphology and distribution of red-tide dinoflagellate cysts in China[D]. Dalian: Dalian Maritime University, 2016. [15] 黄海燕, 陆斗定, 夏平, 等. 2006年冬季长江口海域表层沉积物中甲藻包囊的分类学研究[J]. 生态学报, 2009, 29(11): 5902−5911. doi: 10.3321/j.issn:1000-0933.2009.11.019Huang Haiyan, Lu Douding, Xia Ping, et al. Taxonomic study of dinoflagellate cysts in Changjiang Estuary in the winter of 2006[J]. Acta Ecologica Sinica, 2009, 29(11): 5902−5911. doi: 10.3321/j.issn:1000-0933.2009.11.019 [16] 黄琳. 东、黄海海域沉积物中甲藻包囊的生态分布特征及其与海区污染关系研究[D]. 广州: 暨南大学, 2012.Huang Lin. Ecological distribution of dinoflagellate cysts in the sediments of the East China Sea and Yellow Sea and its relationships with marine pollution[D]. Guangzhou: Jinan University, 2012. [17] Blackburn S I, Hallegraeff G M, Bolch C J. Vegetative reproduction and sexual life cycle of the toxic dinoflagellate Gymnodinium catenatum from Tasmania, Australia[J]. Journal of Phycology, 1989, 25(3): 577−590. doi: 10.1111/j.1529-8817.1989.tb00264.x [18] 许翠娅. 泉州湾春、夏季浮游植物的生态特征[J]. 福建水产, 2001(4): 64−69.Xu Cuiya. Ecological characteristics of phytoplankton in Quanzhou Bay in spring and summer[J]. Journal of Fujian Fisheries, 2001(4): 64−69. [19] 江兴龙, 宋立荣. 泉州湾赤潮藻类优势种演替影响因子探讨[J]. 海洋与湖沼, 2009, 40(6): 761−767. doi: 10.3321/j.issn:0029-814X.2009.06.014Jiang Xinglong, Song Lirong. The influence factors on dominant red-tide algal species succession in Quanzhou Bay[J]. Oceanologia et Limnologia Sinica, 2009, 40(6): 761−767. doi: 10.3321/j.issn:0029-814X.2009.06.014 [20] 李涛, 刘胜, 黄良民, 等. 大亚湾一次赤潮生消期间浮游植物群落变化研究[J]. 热带海洋学报, 2005, 24(3): 18−24. doi: 10.3969/j.issn.1009-5470.2005.03.003Li Tao, Liu Sheng, Huang Liangmin, et al. Studies on phytoplankton community change at Daya Bay during a red tide[J]. Journal of Tropical Oceanography, 2005, 24(3): 18−24. doi: 10.3969/j.issn.1009-5470.2005.03.003 [21] 庞勇, 聂瑞, 吕松辉. 珠江口双胞旋沟藻Cochlodinium geminatum赤潮生消过程的环境特征初步分析[J]. 生态环境学报, 2015, 24(2): 286−293.Pang Yong, Nie Rui, Lü Songhui. Preliminary environmental analysis of the evolution of Cochlodinium geminatum bloom in the Pearl River Estuary[J]. Ecology and Environmental Sciences, 2015, 24(2): 286−293. [22] 邓华, 管卫兵, 曹振轶, 等. 2012年福建沿海大规模米氏凯伦藻赤潮暴发的水文气象原因探讨[J]. 海洋学研究, 2016, 34(4): 28−38. doi: 10.3969/j.issn.1001-909X.2016.04.004Deng Hua, Guan Weibing, Cao Zhenyi, et al. Analysis of hydrological and meteorological factors causing Karenia mikimotoi bloom in 2012 along Fujian coast[J]. Journal of Marine Sciences, 2016, 34(4): 28−38. doi: 10.3969/j.issn.1001-909X.2016.04.004 [23] Liu Minlu, Gu Haifeng, Krock B, et al. Toxic dinoflagellate blooms of Gymnodinium catenatum and their cysts in Taiwan Strait and their relationship to global populations[J]. Harmful Algae, 2020, 97: 101868. doi: 10.1016/j.hal.2020.101868 [24] Zheng Chongrong, Li Guangyi, Chen Jingwu, et al. Potential effects of some environmental factors on a dinoflagellate red tide caused by Gymnodinium catenatum in Shenhu Bay in 2017[J]. Meteorological and Environment Research, 2019, 10(4): 26−30. [25] Hallegraeff G M, Steffensen D A, Wetherbee R. Three estuarine Australian dinoflagellates that can produce paralytic shellfish toxins[J]. Journal of Plankton Research, 1988, 10(3): 533−541. doi: 10.1093/plankt/10.3.533 [26] Figueroa R I, Bravo I, Ramilo I, et al. New life-cycle stages of Gymnodinium catenatum (Dinophyceae): laboratory and field observations[J]. Aquatic Microbial Ecology, 2008, 52(1): 13−23. [27] 程军利, 张鹰, 张东, 等. 海州湾赤潮发生期生态环境要素分析[J]. 海洋科学进展, 2009, 27(2): 217−223. doi: 10.3969/j.issn.1671-6647.2009.02.012Cheng Junli, Zhang Ying, Zhang Dong, et al. Analysis of ecological environment elements during the red tide occurring in Haizhou Bay[J]. Advances in Marine Science, 2009, 27(2): 217−223. doi: 10.3969/j.issn.1671-6647.2009.02.012 [28] 高铭琪. 石狮市近岸海域水质分析与评价[J]. 渔业研究, 2017, 39(6): 493−501.Gao Mingqi. Analysis and evaluation of water quality in Shishi inshore waters[J]. Journal of Fisheries Research, 2017, 39(6): 493−501. [29] 丁光茂, 洪华生, 王大志. 东海原甲藻和链状亚历山大藻对硝酸盐和氨盐的生理响应[J]. 厦门大学学报(自然科学版), 2010, 49(1): 95−101.Ding Guangmao, Hong Huasheng, Wang Dazhi. Physiological response of Prorocentrum donghaiense and Alexandrium tamarense to nitrate and ammonium[J]. Journal of Xiamen University (Natural Science), 2010, 49(1): 95−101. [30] Bravo I, Fraga S, Figueroa R I, et al. Bloom dynamics and life cycle strategies of two toxic dinoflagellates in a coastal upwelling system (NW Iberian Peninsula)[J]. Deep-Sea Research Part II: Topical Studies in Oceanography, 2010, 57(3/4): 222−234. [31] Bolch C J S, Hallegraeff G M. Dinoflagellate cysts in recent marine sediments from Tasmania, Australia[J]. Botanica Marina, 1990, 33(2): 173−192. [32] 潘俊, 李瑞香, 李艳, 等. 秋季南黄海表层沉积物中甲藻包囊分布[J]. 海洋科学进展, 2010, 28(1): 41−49. doi: 10.3969/j.issn.1671-6647.2010.01.006Pan Jun, Li Ruixiang, Li Yan, et al. Distribution of dinoflagellate cysts in surface sediments from the southern Yellow Sea in autumn[J]. Advances in Marine Science, 2010, 28(1): 41−49. doi: 10.3969/j.issn.1671-6647.2010.01.006 [33] 李光毅, 郑崇荣, 吴惠锴, 等. 湄洲湾海域甲藻包囊的时空分布初步调查[J]. 海洋学报, 2020, 42(8): 115−126.Li Guangyi, Zheng Chongrong, Wu Huikai, et al. A preliminary investigation of spatial and temporal distribution of dinoflagellate cysts in the Meizhou Bay[J]. Haiyang Xuebao, 2020, 42(8): 115−126. [34] Figueroa R I, Bravo I, Garcés E, et al. Nuclear features and effect of nutrients on Gymnodinium catenatum (Dinophyceae) sexual stages[J]. Journal of Phycology, 2006, 42(1): 67−77. doi: 10.1111/j.1529-8817.2006.00181.x -
 下载:
下载:
计量
- 文章访问数: 800
- HTML全文浏览量: 304
- PDF下载量: 45
- 被引次数: 0
